|






 |  |

| Biologie | Botanica | Chimie | Didactica | Fizica | Geografie |
| Gradinita | Literatura | Matematica |
Biologie
|
|
Qdidactic » didactica & scoala » biologie Structura si componentele celulei |
Structura si componentele celulei
STRUCTURA SI COMPONENTELE CELULEI
1. MEMBRANA CELULARǍ
Determina forma celulei si are functie protectoare. A fost semnalata pentru prima data de Robert Hooke (1665), dar a fost definita precis de Moldenhauer in 1812.
Membrana celulara isi are originea in procesul de citochineza cand se formeaza lamela mijlocie din fragmoplast care reprezinta punctul de plecare al formarii membranei. O data cu lamela mijlocie se formeaza si plasmodesmele.Peste lamela mijlocie se depun microfibrile de celuloza formandu-se membrana primara. La celulele specializate se intensifica procesul de depunere al celulozei si ligninei, formandu-se membrana secundara. Un rol important in depunerea microfibrilelor de celuloza si lignina il au aparatul Golgi si reticulul endoplasmatic.
Modificarile secundare ale membranei celulozice.
Prin specializarea celulelor la indeplinirea anumitor functii, membrana celulozica isi schimba adesea compozitia, structura si functia. Peste membrana primara se pot depune noi straturi de substante scheletice secretate de protoplast. Asemenea schimbari pe care le sufera membrana celulelor poarta numele de modificari secundare ale membranei celulozice. Modificarile secundare mai frecvent intalnite sunt urmatoarele:
a) Cerificarea se intalneste la celulele epidermice si consta in acoperirea membranei acestora cu un strat de ceara, facandu-le putin permeabile sau impermeabile pentru apa. Se intalneste la mere, pere, struguri, frunzele de conifere etc.
b) Mineralizarea consta in incrustarea (impregnarea) membranei celulare cu substante minerale. Mai des intalnim incrustarea cu SiO2 la tulpina de grau si coada calului, si cu CaCO3, la Corallina si Chara.
c) Cutinizarea consta in impregnarea membranei celulelor epidermice cu o substanta de natura grasa numita cutina. Stratul continuu de cutina de la exteriorul organelor vegetative poarta numele de cuticula. Cutina amestecata cu ceara devine impermeabila pentru apa (in cazul unor xerofite).
d) Lignificarea se intalneste numai la plantele superioare si consta in impregnarea membranei celulozice cu lignina. Se observa la vasele lemnoase, fibrele lemnoase si fibrele de sclerenchim.
e) Suberificarea consta in impregnarea intregii membrane celulozice cu suberina. Suberina este de natura grasa impermeabila pentru apa si gaze, fapt care face ca celulele cu membranele suberificate sa moara.
f) Gelificarea consta in impregnarea membranei cu substante pectice care in contact cu apa se umfla si se transforma in gume sau mucilagii.
g) Lichefierea este fenomenul dizolvarii si disparitiei complete a membranei celulare. Se intalneste la formarea traheelor, laticiferelor si spatiilor intercelulare.
h) Taninizarea consta in impregnarea membranei celulozice cu taninuri fapt care face ca aceste celule sa devina imputrescibile pe o perioada indelungata de timp.
2. CITOPLASMA
La procariote se caracterizeaza printr-o stare coloidala de gel permanent, stare care exclude existenta curentilor citoplasmatici si asigura mentinerea structurii nucleului care este lipsit de membrana limitanta.La eucariote citoplasma prezinta mobilitate. Citoplasma fundamentala (hialoplasma) prezinta proprietati fizico-chimice asemanatoare cu ale plasmalemei.Celulele eucariote au capacitatea de a adaposti simbionti de tip procariot sau eucariot ca de exemplu bacterii din genul Rhizobium in celulele radiculare ale nodozitatilor de la leguminoase.
Organitele celulare:
In citoplasma celulelor eucariote vegetale sunt localizate organite celulare cu functii specifice ca: nucleul, reticulul endoplasmatic, mitocondriile, ribozomii, plastidele, aparatul Golgi s.a., dintre care numai ribozomii se intalnesc si in celula procariota, celelalte fiind absente sau difera structural de cele din celula eucariota.
RETICULUL ENDOPLASMATIC
A fost observat in 1945 de Porter si Fulam. in anul 1954, Palade si Porter au constatat ca reticulul endoplasmatic este reprezentat printr-un sistem de interconectare a canalelor, veziculelor si a cisternelor ultramicroscopice. Unele portiuni ale reticulului endoplasmatic au suprafata rugoasa, fiind tapisate la exterior cu ribozomi, iar altele netede, ribozomii lipsind aici.
Functiile reticulului endoplasmatic:
- Participa la transportul substantelor in citoplasma si la eliminarea unora in mediul extracelular.
- Participa la transportul substantelor nutritive in corpul plantelor, deoarece formatiile canaliculare ale reticulului endoplasmatic sunt orientate in directia circuitului de seva elaborata.
- Vasilieu (1969) arata ca reticulul endoplasmatic rugos are rol in sinteza substantelor proteice care apoi vor intra in structura reticulului endoplasmatic neted. De asemenea mai are rol si in sinteza unor enzime.- in cisternele reticulului endoplasmatic, situate langa vacuole, se formeaza substante proteice care apoi trec in vacuole, mecanismul de trecere ramanand insa necunoscut.
- Se pare ca membranele reticulului endoplasmatic participa la sinteza substantelor necesare membranei celulozice si membranei lignificate.
- Mathile si Moor (1968) considera ca din elementele reticulului endoplasmatic se formeaza niste vezicule sau provacuole care reprezinta de fapt lizozomi, apoi mai multe provacuole se unesc si formeaza vacuole adevarate.
- Se pare ca atunci cand functia mitocondriei inceteaza, ea este preluata temporar de reticulul endoplasmatic.
VACUOLA
Vacuola este un organit important al celulei vegetale, avand un suc celular si fiind marginita de o membrana plasmatica numita tonoplast.
APARATUL GOLGI
Incepand cu anul 1957, Buvat si Porter il descriu in celula vegetala. Totalitatea dictiozomilor dintr-o celula formeaza aparatul Golgi. La microscopul electronic s-a observat ca dictiozomul este o formatiune lamelara sub forma de cisterne suprapuse unele peste altele, unite prin partea lor centrala, cu marginile ramanand libere. O cisterna este formata din doua membrane elementare sudate la capete, iar intre ele se afla un spatiu plin cu lichid transparent. De la marginea libera a cisternelor se detaseaza mereu vezicule sferice ce migreaza in citoplasma. De obicei membranele dictiozomilor nu au ribozomi; la Caulerpa poarta ribozomi insa.
LIZOZOMII
In anul 1949 De Duve ii izoleaza din celulele ficatului de
sobolan. La plante se vorbeste de lizozomi incepand cu anul 1965 cand
Matille si colab. arata ca sferozomii si lizozomii sunt
unul si acelasi lucru din punct de vedere ontogenetic. De asemenea la
Fungi sferozomii functioneaza si ca lizozomi. Din aceasta cauza
RIBOZOMII
Printre primii care-i observa in celula animala este G. Palade. La plante, Robinson si Brown in 1953 ii observa in celulele radacinii de bob. Ribozomii sunt distribuiti in citoplasma, nucleu, plastide si mitocondrii. in citoplasma pot fi liberi sau fixati de reticulul endoplasmatic. Ribozomii cloroplastelor si mitocondriilor sunt mai mici, iar cei mitocondriali au un ARN specific.
Ribozomii sunt formati din 40-50% ARN, 50-60% proteine si foarte putine lipide. Referitor la geneza ribozomilor exista mai multe pareri. Unii citologi sustin formarea ribozomilor la nivelul nucleolului, altii sustin autoreproducerea lor, iar altii sustin formarea lor de novo din plasma de baza. Cercetarile facute cu microscopul electronic pledeaza pentru originea nucleolara a ribozomilor, iar de aici migreaza in citoplasma prin porii invelisului nuclear. Pana in prezent nu se stie daca ribozomii plastidiali si mitocondriali se formeaza tot in nucleoli sau isi au originea chiar in cloroplaste si mitocondrii.
Ribozomii au rol important in sinteza proteinelor.
PLASTIDELE
In anul 1883 Schimper a descris trei tipuri de plastide tinand cont de culoarea lor: leucoplaste, cromoplaste si cloroplaste, considerand ca tip de origine leucoplastele.Totalitatea plastidelor dintr-o celula formeaza plastidomul celular.
Leucoplastele sunt plastide incolore. Se pot afla sub forma de amiloplaste, elaioplaste si proteinoplaste. Forma lor este sferica sau eliptica, dar isi modifica aspectul atunci cand in ele se formeaza grauncioare de amidon sau cristale proteice. Ca si proplastidele, in prezenta luminii, sunt capabile sa formeze structuri tilacoidale si sa capete culoarea verde.
Cloroplastele sunt plastidele verzi care se gasesc in toate organele verzi ale plantelor. Forma si marimea lor este variabila. Ele executa o miscare pasiva in celula fiind antrenate de curentii citoplasmatici.
CENTROZOMUL (CENTRUL CELULAR)
Este situat in apropierea nucleului. A fost pus in evidenta numai in celulele plantelor inferioare si la animale. Ca structura el este alcatuit din centriol, microcentru (centrozom), centrosfera si astrosfera. In profaza centrul celular se divide, iar cele doua jumatati migreaza la doi poli opusi ai celulei, in metafaza filamentele celor doi centrozomi se alungesc spre centrul celulei formand fusul de diviziune. Se pare ca si celulele plantelor superioare ar avea un centrozom care s-ar numi centru primar de atractie, iar dupa ce se divide si migreaza la cei doi poli ai celulei s-ar numi centrii polari de atractie (Gimnez-Martin si colab., 1962). Cu privire la originea centrului celular cele mai multe pareri considera ca se autoreproduc printr-un proces de diviziune, dublandu-se numarul lor.
3. NUCLEUL
A fost descoperit de R. Brown in anul 1831 in celulele
orhideelor. La unele bacterii si alge albastre nu s-a evidentiat un
nucleu morfologic. Organismele care nu au nucleu distinct au fost numite
procariote, iar cele cu nucleu distinct eucariote. Forma si marimea
nucleului este variata in lumea vegetala, depinzand de tipul de
celule. Ca structura nucleul prezinta la exterior un invelis
dublu care lasa din loc in loc
Din punct de vedere chimic nucleul este format din ADN asociat cu histone, proteine acide, ARN si fosfolipide. Proprietatile fizice ale nucleului difera de cele ale citoplasmei. Miscarile ameoboidale ale nucleului si capacitatea nucleului de a fuziona in timpul fecundatiei ne arata starea de sol a nucleului. Faptul ca masa nucleara se poate intinde in fire subtiri cu ajutorul micromanipulatorului si ca uneori nucleii se alatura fara a fuziona demonstreaza starea de gel a nucleului.
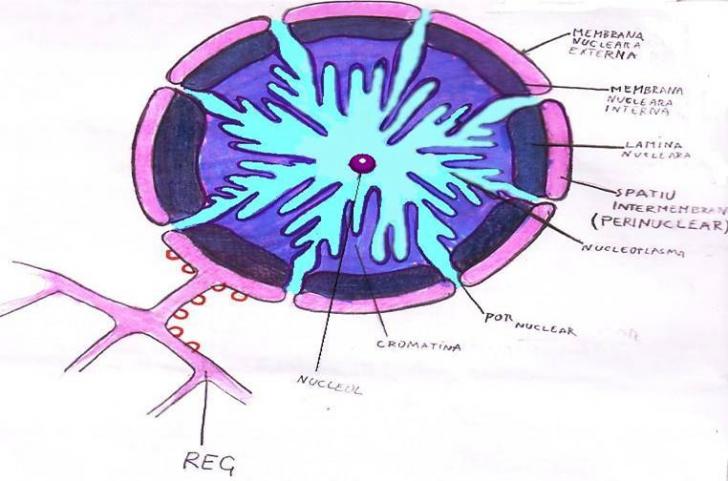
MEMBRANA NUCLEARA
Invelisul nuclear este dublu, de tip elementar, iar intre cele doua membrane se afla spatiul perinuclear. In acest spatiu se gaseste un lichid cu densitate mica ce seamana cu un fel de ser. Se pare ca acest lichid este asemanator cu cel din reticulul endoplasmatic care poarta numele de enchilem. Invelisul nuclear nu este numai o bariera mecanica intre masa nucleara si citoplasma, ci reprezinta o formatiune vie cu multiple roluri in viata celulelor.
CARIOPLASMA
Este constituita dintr-un lichid transparent si omogen, care contine un amestec de substante proteice. El reprezinta mediul in care au loc toate transformarile cromozomilor.
NUCLEOLUL
Este dispus in masa nucleului si este partea cea mai densa a acestuia. Poate fi sferic, eliptic, alungit sau sinusoidal. El nu prezinta o membrana proprie care sa-l izoleze de restul masei nucleare. Volumul si numarul nucleolilor dintr-un nucleu variaza in functie de starea fiziologica a celulei, de conditiile de mediu, de intensitatea sintezei substantelor proteice etc. Numarul nucleolilor poate fi de 1-3, la unele alge ajungand pana la 100. Masa de baza a nucleolului este formata din proteine bazice si fosfolipide legate de ARN, in nucleol nu s-a evidentiat ARN informational. In celula vegetala nucleolul indeplineste mai multe functii:
1. Rolul nucleolului in diviziunea celulara. Unii presupun ca
substanta nucleolara ia parte la formarea structurilor fusoriale,
si alti cercetatori arata ca, datorita
continutului bogat in ribonucleoproteine, nucleolul participa la
schimburile de substante.
2. Rolul nucleolului in sinteza
proteinelor si a ARN. S-a demonstrat prin metode autoradiografice, ca
nucleolul are unele posibilitati de sinteza a ARNului insa
marea majoritate a ARN-ului nucleolar se sintetizeaza in cromatina.
De asemenea, nucleolul este capabil sa sintetizeze substante
proteice, in special histone.
|
Structura celulei vegetale

4. CROMOZOMII
Transmiterea caracterelor de la parinti la urmasi a constituit, din cele mai vechi timpuri, unul dintre subiectele de studiu si analiza ale omenirii. Dupa vizualizarea la microscopul optic a nucleului celular si stabilirea importantei sale prin observarea fuziunii gametilor in procesul de fertilizare, a venit randul descoperirii rolului cromozomilor, ca si constituenti nucleari, in procesul transmiterii caracterelor.
Denumite "corpuri colorate" ( din grecescul croma = culoare si soma = corp) datorita vizualizarii lor la microscopul optic in urma colorarii lor cu coloranti bazici, cromozomii, s-a constatat ca poseda trei caracteristici importante:
Nucleul oricarei celule somatice prezinta un numar fix de cromozomi, tipic pentru o anumita specie, ce poarta denumirea de complet cromozomal (2n). Numarul cromozomilor variaza foarte mult de la o specie la alta si nu prezinta nici o legatura cu gradul de complexitate al organismului (tabelul nr. 2.1).
|
Specia la plante |
2n |
Specia la animale |
2n |
|
Triticum monococcum |
|
Homo sapiens sapiens |
|
|
Triticum durum |
|
Equus caballus |
|
|
Triticum aestivum |
|
Bos Taurus |
|
|
Secale cereale |
|
Sus scrofa |
|
|
Hordeum vulgare |
|
Ovies aries |
|
|
Zea mays |
|
Canis familiaris |
|
|
Solanum tuberosum |
|
Mus musculus |
|
|
Helianthus annuus |
|
Cavia cobaya |
|
|
Avena sativa |
|
Oryctolagus cuniculus |
|
|
Brassica oleracea |
|
Gllus domesticus |
|
|
Beta vulgaris |
|
Rana esculenta |
|
|
Glycine max |
|
Felix domestica |
|
|
Medicago sativa |
|
Musca domestica |
|
|
Nicotiana tabacum |
|
Drosophila melanogaster |
|
|
Oryza sativa |
|
Culex pipiens |
|
|
Phaseolus vulgaris |
|
Apis mellifica |
|
|
Pisum sativum |
|
Euphragurus eshotensis |
|
|
Trifolium pretense |
|
Locusta migratoria |
|
|
Allium cepa |
|
Bombyx mori |
|
|
Capsicum anuum |
|
Cyprinus carpio |
|
|
Malus silvestris |
|
Lepus cuniculus |
|
|
Prunus domestica |
|
Vulpes vulpes |
|
|
Pirus communis |
|
Phyloxera caryaecaulis |
|
|
Vitis vinifera |
|
|
|
2.Cromozomii din cadrul completului cromozomal sunt, de regula, prezenti in perechi. De exemplu, completul cromozomal al mazarei contine 14 cromozomi, deci 7 perechi. In cadrul perechii, unul dintre cromozomi este de origine materna, iar celalalt de origine paterna. Atunci cand nucleul contine perechi de cate doi cromozomi similari, unul matern si celalalt patern, spunem ca este diploid, iar organismul, la randul sau, este diploid. Daca exista trei cromozomi similari, avem de a face cu un nucleu triploid, daca sunt patru, nucleul este tertraploid, s.a.m.d.
3. Celulele germinale, sau gametii, spre deosebire de celulele somatice, contin doar unul din membrii perechii, deci jumatate din numarul de cromozomi ai completului cromozomal si se numesc haploide (n). Aceasta stare de haploidie este necesara deoarece, in momentul unirii gametilor prin procesul de fertilizare, se reface starea diploida caracteristica celulei zigot.
Faptul ca organismele complexe rezulta din unirea a doua celule haploide si ca, apoi, prezinta celule somatice diploide si celule gametice haploide, indica prezenta a doua procese de diviziune nucleara. Unul dintre ele - numit mitoza - mentine intact numarul de cromozomi din nucleul celulelor, in timp ce al doilea - numit meioza - reduce la jumatate numarul acestora.
4. 1. STRUCTURA CROMOZOMILOR
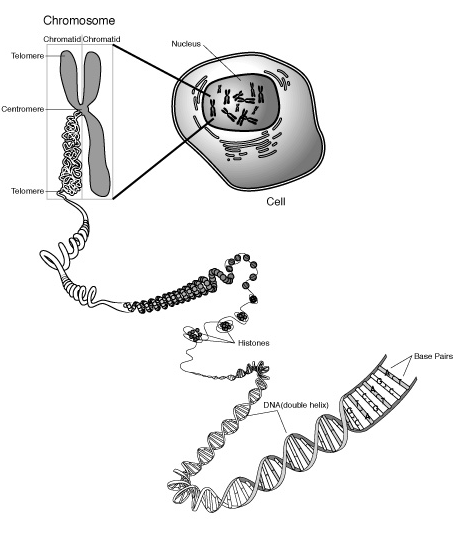
Structura cromozomilor devine vizibila la microscop in momentul de maxima impachetare a cromatinei, in metafaza mitotica. Fiecare cromozom poseda doua unitati structurale identice denumite cromatide unite la nivelul centromerului ( constrictia primara ). La randul sau, fiecare cromatida este alcatuita din doua filamente subtiri, denumite cromoneme, care in metafaza, sufera un proces puternic de spiralizare, ducand la scurtarea si ingrosarea cromozomilor. Datorita spiralizarii si rotirii cromonemelor, se formeaza corpusculi cromatici, denumiti cromomere.
Fig. 2. 3. Structura fizica a cromozomului

Centromerul este resposanbil pentru miscarea cromozomilor in mitoza si meioza, asigurand atasarea acestora de fibrele fusului de diviziune. El reprezinta centrul mobil si viabil al cromozomului. Fara el, cromozomul nu este viabil si se resoarbe. In mod normal, cromozomii prezinta un singur centromer. Datorita unor dislocatii pot apare cromozomi cu doi sau mai multi centromeri ( dicentrici sau policentrici ) sau cromozom fara centromer ( acentrici ). Unii cromozomi pot prezenta, pe langa constrictia primara si o constrictie secundara sau organizator nucleolar, portiunea de cromozom ramasa dupa aceasta purtand numele de satelit. Capetele cromozomilor, denumite telomere nu permit atasarea sau pierderea unor segmente de cromozom, asigurand astfel stabilirea si integritatea acestora.
Cromozomii, vizibil in metafaza, pot fi identificati pe seama caracteristicilor lor morfologice stabile pe parcursul diviziunilor mitotice si meiotice. Astfel, fiecare dintre ei poseda o anumita forma, o anumita marime, anumite particularitati. Pe baza caracteristicilor lor morfologice sunt alcatuite cariotipurile speciilor, ajutand astfel la identificarea si diferentierea lor.
In general, forma cromozomilor este data de pozitia centromerului pe cromozom. Un cromozom avand centromerul in mijloc, este numit metacentric; el are bratele egale, iar in timpul anafazei are forma literei V. Cromozomul cu centromerul situat in apropierea de unul din capete este numit submetacentric si are forma literei J. in anafaza, iar cel cu centromerul aflat la unul din capete este acrocentric si are forma literei I.
Idiograma la om

4. 2. COMPOZITIA CHIMICǍ A CROMOZOMILOR
Componenta principala a cromozomilor este reprezentata de acidul dezoxiribonucleic (ADN ). Acesta, in cazul eucariotelor este impachetat si asociat cu molecule proteice in agregate stabile numite cromatina. Alaturi de acestea, in compozitia cromozomilor, mai intra si un procent redus (5%) de acid ribonucleic (ARN ). Clasa principala de proteine componente ale cromatinei o reprezinta histonele.
Fig. 2. 4. Stadii de condensare ale cromozomului: 1- ADN; 2 - cromatina (ADN + histone); 3 - cromatina pe parcursul interfazei; 4 - cromatina pe parcursul profazei; 5 - cromatina pe parcursul metafezei.

Cinci mari tipuri de histone sunt prezente in componenta cromozomilor : H1, H2A, H2B, H3 si H4. Histonele, fiind incarcate cu sarcini pozitive, adera la ADN-ul incarcat negativ. De asemenea, acestea se leaga intre ele, toate legaturile proteine -ADN, proteine-proteine asigurand structura cromatinei. Unitatea de baza a cromatinei o reprezinta nucleosomul alcatuit din: o molecula de H1si H4 cate doua molecule de H2A, H2B, H3 si un segment de ADN de aproximativ 200 de nucleotide. Aranjarea cromatinei in cromozom nu este complet cunoscuta, insa se stie ca aceasta este impachetata din ce in ce mai mult, astfel incat in metafaza mitotica ajunge la gradul cel mai inalt de impachetare fapt ce permite vizualizarea cromozomilor la microscopul optic.
In zona centromerului, a organizatorului nucleolar si a telomerelor, cromozomul apare mai intens colorat, fiind alcatuit din heterocromatina. Restul cromozomului este alcatuit din eurocromatina. Cele doua tipuri de cromatina poseda proprietati biochimice si functii genetice diferite.
Fig. 2. 5. Nucleosomul

4. 3. TIPURI PARTICULARE DE CROMOZOMI
Cercetarile de citogenetica au scos in evidenta ca, la anumite specii, exista, pe langa cromozomii obsnuiti, si anumite tipuri particulare de cromozomi. Acestia au marimi, forme si caracteristici genetice diferite fata de cromozomii normali.
Cromozomii politeni, identificati in celulele glandelor salivare de la larvele de Drosophila melanogaster sunt cromozomi gigantici, de cateva ori mai mari decat cromozomii speciei. Acestia contin aproximativ 1000 de molecule de ADN aliniate una langa cealalta si au rezultat prin duplicarea repetata a ADN-ului fara ca moleculele formate sa se separe. Datorita marimii si morfologiei lor, cromozomii politeni reprezinta un excelent obiect de studiu pentru hibridarea in situ a acizilor nucleici.
Fig. 2. 6. Cromozom politen la Drosophila

Cromozomii suplimentari au fost observati in celulele somatice ale unor specii de plante ( porumb, secara, sorg) ca fiind in plus fata de completul cromozomal al acestora. In plus, ei se coloreaza mai intens si se situeaza la periferia fusului de diviziune. Au activitate genetica slaba cu efecte nu intotdeauna vizibile asupra fenotipului plantei.
Cromozomii perie de lampa au, datorita cromatidelor care formeaza bucle in jurul unui ax comun, aspectul specific periilor de lampa. Sunt foarte subtiri si, prin redresarea buclelor revin la morfologia cromozomilor obisnuiti.
| Contact |- ia legatura cu noi -| | |
| Adauga document |- pune-ti documente online -| | |
| Termeni & conditii de utilizare |- politica de cookies si de confidentialitate -| | |
| Copyright © |- 2024 - Toate drepturile rezervate -| |
|
|
|||
|
|||
|
|||
Proiecte pe aceeasi tema | |||
|
| |||
|
|||
|
|
|||