|






 |  |

| Biologie | Botanica | Chimie | Didactica | Fizica | Geografie |
| Gradinita | Literatura | Matematica |
Biologie
|
|
Qdidactic » didactica & scoala » biologie Teoria cromozomala a ereditatii |
Teoria cromozomala a ereditatii
TEORIA CROMOZOMALA A EREDITATII
Pe baza incrucisarilor la mazare, Mendel a observat si a explicat fenomenul de segregare si combinare independenta a genelor, care afecteaza in egala masura si cromozomii, fiecare gena fiind localizata pe un cromozom. In felul acesta, in meioza, are loc o distributie independenta a fiecarui cromozom din perechea parentala, intr-un gamet sau altul. Intrucat in experientele sale Mendel a folosit la incrucisare indivizi care posedau caractere determinate de gene independente, localizate fiecare pe cate un cromozom, genele dintr-un heterozigot de tipul AaBb s-au separat in meioza independent si s-au combinat in procesul de fecundare liber in combinatii posibile, intr-un raport fenotipic de 9:3:3:1.
In anul 1906, W. Bateson si R.C. Punnett (citati de T.Craciun si colab., 1978) au realizat o dihibridare la mazare, incrucisand un soi cu flori purpurii si polen cilindric cu un alt soi de flori rosii si polen sferic. In prima generatie, F1, au rezultat numai plante cu flori purpurii si graunciori de polen cilindrici, relevand fenomenul de dominanta completa (aceasta a permis simbolizarea parintilor cu AABB si aabb, iar hibridul F1 cu AaBb). Prin autofecundarea plantelor F1 a rezultat generatia F2 cu un raport global de segregare de 7:7:1:1, in care formele parentale (flori purpurii cu polen cilindric si flori rosii cu polen sferic) au aparut cu frecvente mari si egale (7:7), iar combinatiile noi intre acestea au aparut cu frecvente mici si in proportii de asemenea egale (1:1) (flori rosii-polen cilindric si flori purpurii-polen sferic).
Pentru verificarea acestui raport de segregare, care era cu totul diferit fata de raportul mendelian de 9:3:3:1, au fost incrucisate alte soiuri cu aceleasi caractere, dar in combinatii diferite (flori purpurii-polen sferic si flori rosii-polen cilindric). In F1 s-au manifestat aceleasi caractere dominante (flori purpurii si graunciori de polen cilindrici), iar in F2 s-a confirmat raportul de 7:7:1:1, in care formele parentale au aparut in exces (7/16 flori purpurii-polen sferic si 7/16 flori rosii-polen cilindric) si combinatiile noi cu o frecventa mica (1/16 flori purpurii-polen cilindric si 1/16 flori rosii-polen sferic). In toate cazurile, dihibridul din F1 (AaBb) a produs 4 tipuri de gameti din a caror fecundare au rezultat 16 combinatii de gene. Frecventa diferita a gemetilor a modificat raportul de segregare fenotipica. Pentru a explica aceste rezultate "cu totul aparte" autorii acestor experiente au emis teoria "cuplarii" si "repulsiei", in virtutea careia faza de cuplare reprezinta situatia in care cele doua alele dominante si respectiv recesive, de la acelasi parinte tind sa intre in proportii mari in gameti si sa fie transmise impreuna (AABB x aabb), iar faza de repulsie, situatia in care o alela dominanta si una recesiva tind sa intre in gameti in proportii mai mari .

Revine meritul lui Thomas Hunt Morgan (1866-1945) si colaboratorilor sai (C.B. Bridges, A.A. Sturtevant si H.J. Muller) care intre anii 1910 - 1915 au efectuat numeroase incrucisari la Drosophilla melanogaster si pe baza rezultatelor obtinute au elaborat teoria cromozomiala a ereditatii, care in esenta asociaza genele cu cromozomii.
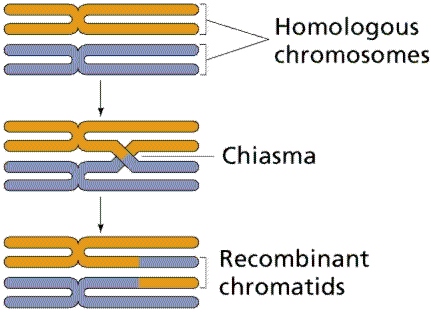
Efectuand cercetari similare pe Drosophilla, ei au demonstrat ca fazele de cuplare si repulsie sunt consecinta localizarii genelor in acelasi cromozom; intre cromozomii omologi poate avea loc un schimb reciproc de gene in proportii diferite, care determina aparitia unor gameti, si respectiv indivizi, recombinati (cu cromozomi recombinati)
Cercetarile si concluziile lui T.H. Morgan si ale colaboratorilor sai au fost sintetizate in trei teze (teorii):

plasarea liniara a genelor pe cromozomi (care a fost asemuita cu aranjarea margelelor intr-un sirag). Aceasta teza s-a bazat pe faptul ca la Drosophilla se cunosteau peste 500 gene mutante, iar in gametii acesteia exista numai 4 cromozomi (2n = 8), si in consecinta, mai multe gene se gasesc in acelasi cromozom;
tendinta genelor de pe acelasi cromozom de a se transmite in bloc sau inlantuite (linkage), adica de a intra in gameti in combinatii parentale, comportandu-se ca o unitate in ereditate. Cand doua sau mai multe gene sunt localizate in acelasi cromozom, spunem ca sunt inlantuite. Ele pot fi inlantuite fie pe un autozom (gene linkage), fie pe un cromozom al sexului (gene sex-likage);
tendinta genelor inlantuite de a intra in gameti in alte combinatii decat cele parentale, acest fenomen fiind denumit crossing-over. In esenta, crossing-over-ul reprezinta un schimb reciproc de segmente cromatidice nesurori intre doi cromozomi omologi. In urma acestui schimb de material genetic (gene) apar cromatide recombinate cu combinatii noi de gene, denumite crossovere sau recombinari, fiind deosebite de combinatiile parentale.
1. 1. INLANTUIREA GENELOR DISPUSE IN ACELASI CROMOZOM (LINKAGE)
Studiul fenomenului de linkage a permis evidentierea mai multor situatii. Astfel, s-a stabilit ca el poate fi complet (absolut sau total), atunci cand genele de pe acelasi cromozom au tendinta permanenta de a ramane inlantuite si incomplet (relativ sau partial), cand intre cromozomii omologi au loc schimburi reciproce de gene.
Detectarea linkage-ului se poate face in doua moduri:
a) prin testcross-ul heterozigotului F1 (mai ales la organismele dioice unde incrucisarea se face usor).
Metoda consta in aparitia in descendenta testcross a unui raport de segregare 1:1, indiferent de numarul de gene inlantuite.
La testcross-ul unui dihibrid heterozigot (F1), in cazul genelor independente (gene localizate pe cromozomi diferiti), rezulta un raport de segregare de 1:1:1:1.
P: AaBb aabb
![]()
![]()
![]()
![]()
![]()
![]()
![]()
![]()
![]() G: AB Ab aB ab x ab
G: AB Ab aB ab x ab
F1: AaBb Aabb aaBb aabb
1 : 1 : 1 : 1
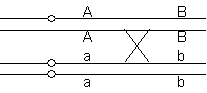
In cazul testcross-ului unui dihibrid heterozigot cu gene inlantuite (situate pe acelasi cromozom) rezulta raportul de segregare de 1:1. reprezentand formele parentale:
P: A_____B ♂ a_____b ♀
____________ ___________
a b a b
![]() G: A_____B a_____b x a_____b
G: A_____B a_____b x a_____b
![]()
![]()
F1: A_____B a_____b
____________ ___________
a b a b
1 : 1
Orice deviere de la raportul testcross de 1:1:1:1 indica o abatere de la combinarea independenta, cel mai adesea datorata fenomenului de linkage.
Exemplu
La incrucisarea unor masculi heterozigoti de drosophilla cu corp cenusiu si aripi normale (BVg/bvg) cu femele dublu recesive cu corp negru si aripi vestigiale (bvg // bvg) folosite ca tester, rezulta in loc de 1 BbVgvg : 1 Bbvgvg : bbVbvb : 1 bbvgvg, cat ar fi rezultat la combinarea indepententa a genelor, un raport simplu de 1 BVg // bvg : 1bvg // bvg, care indica inlantuirea celor doua gene. Acest testcross se prezinta schematic astfel:
P: B_____Vg ♂ b_____vg ♀
_____________ ____________
b vg b vg
G: B_____Vg b_____vg x b_____vg
![]()
![]()
![]()
F1: B_____Vg b_____vg
_____________ ____________
a b b vg
cenusiu, normale negru, vestigiale
: 1
a) prin studiul generatiei F2 (preferabil la organismele hermafrodite si monoice la care testcross-ul se executa mai dificil). In cazul unei dihibridari cu gene independente raportul fenotipic in F2 este de 9:3:3:1, iar la o dihibridare cu gene linkage, precum si la orice polihibridare linkage raportul in F2 este de 3;1. O dihibridare cu gene linkage se desfasoara dupa schema de mai jos.
Linkage-ul complet se manifesta rar. El poate fi prezent numai intr-o pereche sau intr-o regiune oarecare a unei perechi de cromozomi, care nu poate sa conjuge.
cenusiu, normale negru, vestigiale
P: A_____B ♂ a_____b ♀
____________ ___________
a b a b
![]()
![]() G: A_____B x a_____b
G: A_____B x a_____b
F1 A_____B
___________ cenusiu, normale
a b
F2 P: A_____B ♂ A____B ♀
____________ ___________
a b a b
![]()
![]()
![]() G: A_____B a_____b x A_____B a_____b
G: A_____B a_____b x A_____B a_____b
![]()
![]()
![]()
![]()
F2 A_____B A_____B A_____B a_____b
___________ ____________ ___________ __________
A B a b a b a b
|
1 : 2 : 1 -raport
genotipic
3 : 1 - raport fenotipic
1. 2. SCHIMBUL RECIPROC DE GENE (CROSSING-OVER-UL)
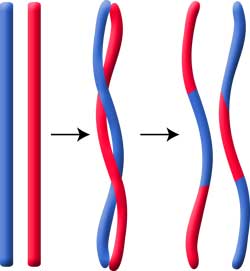
Crossing over-ul apare in meioza, dar poate sa apara si in mitoza, in diferite tesuturi si organe.
In cursul meiozei, fiecare cromozom se duplica in doua cromatide surori, identice. Cromozomii omologi se imperecheaza (conjuga) si formeaza tetrade cromatidice; intre cromatidele nesurori pot avea loc schimburi reciproce de segmente (gene). In schema de mai jos prezentam situatia cand are loc un singur crossing over intre doua gene, A si B.
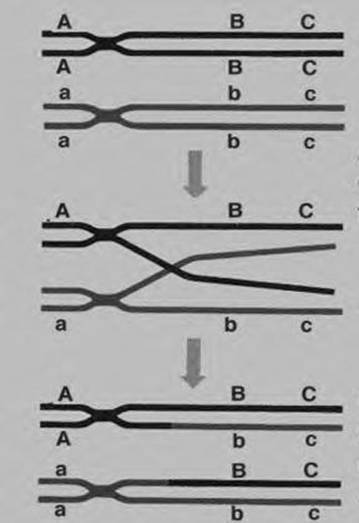
Rezulta ca doua dintre cele patru cromatide nu au suferit crossing over si au genele asociate ca in cromozomii parentali (cromatide parentale), iar celelalte doua cromatide au suferit crossing over-ul (schimb reciproc de gene) si prezinta asociatii noi de gene (cromatide recombinate sau remaniate).
Un crossing over in afara intervalului A-B nu produce schimbari intre cei doi markeri (A si B).
Daca intre doi loci (A-B) apar doua crossing overe intre aceleasi cromatide surori, cromatidele rezultate la sfarsitul meiozei vor fi toate de tip parental.
Orice dublu crossing over intre doi markeri A si B, nu poate fi decelat decat in prezenta unui al treilea marker C situat intre A si B.
Daca intre A si C, pe de o parte si intre C si B, pe de alta parte, probabilitatile de aparitie a unui crossing over sunt x si, respectiv y, probabilitatea crossing overe-lor duble (unul intre A si C si altul intre C si B) este egala cu produsul xy.
Crossing over-ul se detecteaza ca si linkage-ul prin testcross-ul heterozigotului F1 si analiza raportului de segregare in F2.
a) detectarea crossing over-ului prin testcross (la crossing over-ul intre doi loci). Un dublu heterozigot care se testeaza cu parintele dublu recesiv poate avea doua pozitii ale alelelor: i) pozitia cis, cand ambele alele dominante se gasesc pe un cromozom, iar alelele recesive pe cromozomul omolog (AB // ab) si ii) pozitia trans, cand fiecare cromozom are o alela dominanta si alta alela recesiva (Ab // aB). Aceasta indica faptul ca gametii de aceiasi constitutie pot fi parentali sau recombinati dupa asocierea genelor parentale.
Analizele hibridologice efectuate la Drosophila de catre Morgan si colaboratorii au relevat faptul ca heterozigotul F1 folosit ca femela formeaza patru tipuri de gameti in proportii diferite: gametii de tip parental apar cu o frecventa mare, iar gametii recombinati cu o frecventa scazuta. Testerul mascul este homozigot si recesiv, formand un singur tip de gameti. In descendenta testcross apar patru fenotipuri in proportii determinate de cele patru tipuri de gameti. Combinatiile parentale (non cross overe), care apar in proportia cea mai mare, indica intensitatea linkage-ului intre cei doi loci considerati, iar tipurile noi de indivizi reprezinta recombinarile genelor (crossovere). Crossing over-ul se poate prezenta schematic astfel:
P: (cis) A B a b
![]()
![]()
![]()
![]() x ♀ ♂
x ♀ ♂
a b a b
A_____b
a_____B
G: A_____B a_____b x a_____b
![]()
F1: A_____B a_____B A_____B a_____B
____________ ___________ ___________ __________
a b a b a b a b
Combinatii parentale Recombinari
(non crossovere) (crossovere)
P: (trans) A b a b
![]()
![]()
![]()
![]() x ♀ ♂
x ♀ ♂
a B a b
A_____B
a_____b
G: A_____b a_____B x a_____b

![]()
F1: A_____b a_____B A_____B a_____b
____________ ___________ ___________ __________
a b a b a b a b
Combinatii parentale Recombinari intracromozomale
(non crossovere) (crossovere)
Exemplu
La incrucisarea unei femele heterozigote de Drosophila cu corp cenusiu si aripi normale (BVg // bvg0 cu un mascul dublu recesiv (bvg // bvg) au rezultat in descendenta acestui testcross 41,5% indivizi cu corp cenusiu si aripi normale, 41,5% cu corp cenusiu si aripi vestigiale, 8,5% cu corp negru si aripi vestigiale si 8,5% cu corp negru si aripi normale. Primele doua fenotipuri reprezinta combinatiile parentale (non crossovere) care insumeaza 83%, iar ultimele doua fenotipuri reprezinta recombinari ale celor doua gene (crossovere), datorate schimbului reciproc de gene intre cei doi cromozomi omologi, insumand 17%.
Analiza acestor rezultate i-au permis lui Morgan sa constate ca frecventa crossing over-ului intre cele doua gene (B si Vg) reprezinta distanta dintre aceste gene localizate in acelasi cromozom. Cand s-au folosit in analize hibridologice alti loci (gene), frecventa recombinarilor a fost alta, demonstrand astfel, corelatia dintre frecventa crossing overe-lor inregistrate si distanta intre genele implicate.
P: (cis) B____Vg ♀ b____vg ♂
____________ ___________
b vg b vg
B____vg b____vg
G: B____Vg b_ __vg x b____vg
![]()

F1: B____Vg b____vg B____vg b_____Vg
____________ ___________ ___________ ____________
a b a b b vg b vg
Combinatii parentale Recombinari
(non crossovere) (crossovere)
83% 17%
La incrucisarea trans s-a constatat ca intre cei doi loci apare crossing over-ul cu aceiasi frecventa, aceasta constituind un argument in plus pentru dependenta dintre frecventa crossing overe-lor si distanta dintre gene.
b) detectarea crossing over-ului prin analiza descendentei F2 (la crossing over-ul inre cei doi loci). La plante si indeosebi la cele hermafrodite si unisexuat monoice, la care prin testarea heterozigotului s-ar obtine putine seminte (respectiv indivizi) este mai indicata detectarea crossing over-ului prin analiza raportului de segregare in F2.
In cazul a doi loci independenti (nu manifesta linkage), segregarea fenotipica in F2 este tipic mendeliana, adica de 9:3:3:1. Daca cei doi loci manifesta linkage, atunci in F2 se vor manifesta fenotipic in exces combinatiile parentale (non crossoverele), in timp ce recombinarile (crossoverele) vor apare cu o frecventa scazuta, dependenta de distanta dintre genele considerate.
Pentru determinarea gradului de inlantuire (linkage), precum si a frecventei de crossovere, respectiv a distantei dintre gene se pot folosi urmatoarele metode (S. William, 1977):
a) masurarea linkage-ului pe baza raporturilor de segregare in F2. In acest caz, se determina direct in F2 numarul indivizilor ce reprezinta combinatiile parentale si al celor recombinati (crossovere). Se exprima in procente fiecare categorie de indivizi, obtinand astfel, gradul de inlantuire (% de non crossovere) si frecventa recombinarii de gene (% de crossovere) care indica distanta intre gene.
Exemplu: La incrucisarea unui soi cu fructe ovale si inflorescenta compusa (os // os) au rezultat in F1 numai plante cu fructe rotunde si inflorescente simple. Prin autopolenizarea plantelor F1 au rezultat in F2 urmatoarele fenotipuri:
- fruct rotund, inflorescenta simpla - 81 plante
- fruct rotund, inflorescenta compusa -18 plante
- fruct oval, inflorescenta simpla - 22 plante
- fruct oval, inflorescenta compusa -79 plante
Rezulta:
- combinatii parentale: 81 + 79 = 160
- recombinari: 18 + 22 = 40
Total 200
Fercventa combinatiilor (gradul de linkage) este egala cu 160/200 = 0,80 sau 80%. Frecventa recombinarilor (crossoverelor) este egala cu 40/200 = 0,20 sau 20% indicand faptul ca intre cele doua gene O si S exista o distanta de 20 unitati.
b) metoda raportului dintre produsele fenotipurilor recombinate si cele parentale. Se poate calcula frecventa gametilor recombinati a dublu heterozigotilor F1 cu ajutorul frecventelor a, b, c si d a fenotipurilor O. S., O.ss, ooS., ooss aparute in F2. Raportul dintre fenotipurile recombinate si cele parentale se numeste raportul produselor (notat cu x), care este in functie de frecventa de recombinare, ce indica distanta dintre gene.
- dublu heterozigot in cis: x = bc/ad
- dublu heterozigot in trans: x = ad/bc
Valoarea frecventei de recombinare se obtine direct din tabelul 1. cu ajutorul produselor (x).
Tabelul 1.
Frecventa recombinarilor in functie de valoarea raportului produselor (dupa F. R. Immer si M. T. Henderson, 1943)
|
Frecventa recombin. |
Raportul produselor |
Frecventa recombin. |
Raportul produselor |
||
|
ad/bc (trans) |
bc/ad (cis) |
ad/bc (trans) |
bc/ad (cis) |
||
|
|
|
|
|
|
|
Exemplu
Reluand incrucisarea la tomate prezentata in exemplul anterior rezulta ca:
incrucisarea cis
P: OS // OS x os // os
F1: OS // os
F2: Fenotipuri Numar indivizi
(a) O.S. 81
(b) O.Ss 18
(c) ooS. 22
(d) ooss 79
X = bc / ad = 18 x 22 / 81 x 79 = 0,062
In coloana cis din tabel la valoarea X = 0,062 corespunde o frecventa a recombinarilor de circa 19%, foarte apropiata de cea obtinuta prin metoda precedenta.
incrucisarea trans
P: Os // Os x oS // oS
F1: Os // oS
F2: Fenotipuri Numar indivizi
(a) O.S. 20
(b) O.ss 80
(c) ooS. 76
(d) ooss 22
X = ad / bc = 20 x 22 / 80 x 76 = 0,072
In coloana trans din tabel la valoarea X = 0,072 corespunde o frecventa a recombinarilor de circa 19%, foarte apropiata de cea obtinuta prin metoda precedenta.
2. FACTORII CARE INFLUENTEAZǍ
CROSSING - OVER-UL
Daca mediul de viata in care se executa experimentul pentru relevarea crossing over-ului ramane constant de la o generatie la alta, atunci frecventa crossoverelor dintre gene nu se modifica, ceea ce indica faptul ca acest fenomen este determinat genetic. In conditii de mediu cu factori variabili, valoarea crossing over-ului se poate modifica intr-un sens sau altul. De asemenea, anumiti factori biologici sau genetici pot afecta, in anumite cazuri, in mod drastic, valoarea crossing over-ului. De aceea, incrucisarile efectuate in scopul estimarii valorii crossing over se fac in conditii de mediu constante (uniforme) si cu genitori puri.
Dintre factorii care influenteaza direct sau indirect fenomenul de crossing over mentionam pe cei mai semnificativi, si anume (T. Craciun si colab., 1978):
Sexul. La origanismele cu lipsa de omologie intre cromozomii sexului X si Y, insotita adesea de absenta chiasmelor intre autozomii omologi (in profaza I a meiozei), nu apare sau este foarte redus fenomenul de crossing over. De exemplu, la diptere, la care masculul este heterogametic (XY), precum si la unele lepidoptere la care femela este heterogametica (XY), nu apare crossing over-ul. La organismele cu crossing over la ambele sexe, exista adesea diferente de la un sex la altul. De regula, o frecventa mai scazuta a crossoverelor se observa tot la sexul heterogametic.
Varsta. La Drosophila melanogaster, Bridges (1927) a constatat ca femele de varsta diferite produc crossovere in proportii variabile. Astfel, la inceputul maturitatii sexuale se inregistreaza o frecventa maxima de crossovere, iar dupa aceea la intervale diferite de timp, s-au observat mai multe minime. La porumb, s-au observat, de asemenea, fluctuatii ale frecventei de crossovere in raport cu varsta plantelor, iar la femeile intre 35-40 de ani creste rata non-disjunctiei perechii de cromozomi X, ceea ce determina apariti unor descendenti aneuplozi, adesea cu afectiuni (sindromuri) foarte grave.
Zona heterocromatinei
In zona heterocromatinei a cromozomului, situata de o parte si de alta a centromerului, precum si in sateliti, chiasmele lipsesc sau se gasesc cu o frecventa redusa. Genele din aceasta parte a cromozomului sunt foarte dense, puternic spiralizate, cu posibilitati mici de transcriptie si deci, de functionare, fiind considerate inactive.
Modificarile in structura cromozomului
In functie de natura lor, pot reduce sau suprima prezenta chiasmelor intre cromozomii omologi. Aceste modificari apar cu frecventa mare in urma tratamentelor cu agenti mutageni in doze mari, care induc numeroase si diferite dislocatii cu efecte negative asupra organismelor.
Practic, toate tipurile de modificari in structura cromozomilor (deficientele, duplicatiile, inversiile, translocatiile) in stare heterozigota suprima sau reduc aparitia chiasmelor prin lipsa omologiei induse artificial intre cromozomii pereche.
Modificarile numarului de genomuri sau de cromozomi
(autopoliploidia, allopoliploidia, monosomia) reduc sau suprima total aparitia chiasmelor si respectiv, a crossing over-ului.
Factorii de mediu (temperatura, lumina, umididatea, nutritia, etc.) pot afecta in mod diferit, in anumite etape ale dezvoltarii oraganismelor, aparitia chiasmelor si respectiv, a crossing overe-lor, mai ales in regiunile heterocromatice ale cromozomilor.
3. TIPURI DE CROSSING - OVER
Crossing over-ul se poate clasifica dupa mai multe criterii, dintre care mentionam:
a) dupa tipul de diviziune celulara in care apare se cunosc doua tipuri de crossing over, si anume:
1) crossing over meiotic, are loc in profaza I a meiozei si se identifica in diplonem, prin aparitia chiasmelor intre cromatidele nesurori ale cromozomilor omologi. Este tipul cel mai frecvent de crossing over, care prin recombinarile intracromozomiale, reprezinta o sursa foarte importanta a variabilitatii organismelor. Conditiile majore pentru manifestarea fenotipica a acestui tip de crossing over sunt: prezenta obligatorie a cromozomilor omologi in stare heterozigota si realizarea formatiunilor citologice de tetrada cromatidica.
2) crossing over mitotic sau somatic. Mai sus am precizat faptul ca crossing overe-ul este asociat in mod normal, cu diviziunea meiotica. In anumite conditii si cu o frecventa foarte redusa, un fenomen asemanator poate avea loc si in diviziunea mitotica (in celulele somatice de la plante si animale, la sfarsitul interfazei sau in profaza mitotica, cand cromozomii preche au formate cele 4 fire cromatidice si sunt in stare heterozigota. Acest tip de crossing over a fost denumit crossing over mitotic sau somatic si care se detecteaza fenotipic prin segregarea mitotica a genelor marker pe acelasi individ, determinand aparitia de tesuturi mozaicate (tesuturi normale alaturi de tesuturi recombinate). Crossing over-ul mitotic a fost pus in evidenta la Drosophila melanogaster (C. Stern, 1936); la plante (J.H. taylor, 1958), precum si la numeroase microorganisme (bacterii, ciuperci). In figura urmatoare se prezinta mecanismul uni crossing over mitotic la un individ dublu heterozigot (dupa T. Craciun si colab, 1978).
b) dupa nivelul la care are loc ruperea si schimbul de segmente cromozomiale, crossing over-ul poate fi:
1) crossing over egal, atunci cand chiasmele apar intre loci omologi (la acelasi nivel sau punct) si segmentele cromatidice care se schimba sunt egale (identice); acesta este tipul normal de crossing over, cu ruperi si schimburi simetrice (egale) de segmente cromatidice:
2) crossing over inegal, atunci cand chiasma si, respectiv, schimbul segmentelor cromatidice care se schimba sunt inegale. In cazul crossing over-ului inegal dispare conditia de heterozigotie a cromozomilor omologi. Datorita acestui tip aparte de crossing over, intr-o cromatida remaniata vor apare una sau mai multe gene in dublu exemplar, iar fenomenul se numeste duplicatie, iar in cealalta cromatida vor lipsi genele respective, de unde si denumirea de deficienta (vezi figura de mai jos).
Crossig over-ul inegal si consecintele sale asupra cromatidelor implicate
![]()
![]()
![]()
![]() a a a a
a a a a
![]() b b b b
b b b b
c c e c
d d f d
e e c
f f d
e
f
Sturtevant si Morgan (1923) au observat crossing over-ul inegal la Drosophila, la locusul B, segmentul 16 A din cromozomul X. Ochiul normal (B.) are un singur segment 16 A, in timp ce ochiul Bar (BB) este o duplicatie in tandem cu doua segmente 16 A, iar ochiul dublu Bar (BBB) reprezinta o triplicatie cu trei segmente 16 A. De la ochiul normal (forma rotunda), se ajunge la ochi mici, de forma oblonga - tipul Bar - si la ochi foarte mici - la tipul duble Bar- , relevand in mod evident efectul de dozaj al genelor.
3) crossing over-ul nelegitim, atunci cand chiasma apare intre cromozomi heteromorfici care au portiuni (zone) homoloage sau partial homoloage (la indivizi haploizi sau poliploizi).
c) dupa complexitatea materialului genetic implicat in crossing over distingem doua tipuri:
1) crossing over intergenic (intre gene) in care schimbul reciproc intre cromozomii omologi cuprinde segmente de cromatide (cromozomi) cu una sau mai multe gene. Este tipul obisnuit de crossing over a carui valoare de schimb depinde de distanta dintre gene.
2) crossing over intragenic (in cadrul aceleiasi gene), reprezinta schimburi reciproce intre subunitatile unor gene complexe (loci complecsi), denumite subgene sau pseudoalele. Aceste subgene (pseudoalele) au aparut in cadrul unor gene prin mutatii in diferite zone ale acestora si care sunt alelice prin functiile lor. Subgenele pot fi separate si relevate fenotipic prin crrosing over intragenic, analizand un numar foarte mare de indivizi, si in prezenta unor gene marker langa locusul studiat. Pseudoalele sunt detectate fenotipic in pozitia "trans", cand ele se gasesc plasate diferit pe cei doi cromozomi pereche, in timp ce in pozitia "cis" se manifesta fenotipul salbatic. Cele doua pozitii pentru doua gene pseudoalele a1 si a2 sunt urmatoarele:
a1______+
_____________ - pozitia trans, unde se manifesta fenotipic pseudoalele a1
+ a2
a1______a2
______________ - pozitia cis, unde se manifesta fenotipul salbatic
+ +
Studiul unor gene la Drosophila melanogaster a relevat mai muti loci complexi, cu doua sau mai multe pseudoalele. Astfel, Lewis (1955) a observat crossing over intragenic la locii lozenge, white, forked (in cromozomul X), la locusul star in cromozomul II si la alti loci. Asemenea loci complecsi au fost descoperiti si la alte specii de plante si animale (la bumbac, porumb, soareci etc.).
INTERFERENTǍ SI COINCIDENTǍ
Pe baza unor gene la Drosophila melanogaster, H.J. Muller a descoperit in 1916 fenomenul de interferenta, pe care l-a asociat cu tendinta unui crossing over de a impiedica aparitia unui nou crossing over in apropierea sa, pe o anumita distanta. Astfel, la Drosophila melanogaster aparitia unui crossing over pe acelasi brat al cromozomului inhiba aparitia unui alt crossing over pe o distanta de circa 10 unitati si interferenta acestuia scade progresiv cu cresterea distantei dintre gene. Interferenta nu este eficace peste centromer sau la distante mai mari de 45 unitati Morgan. De asemenea, interferenta difera de la un segment cromozomial la altul, de la un cromozom la altul si de la o specie la alta. Pentru estimarea intensitatii interferentei, H.J. Muller a conceput un model de masurare a acestuia cu ajutorul coincidentei, respectiv al coeficientului de coincidenta (Cc), care reprezinta raportul intre crossing overele duble observate experimental si crossing overele duble prevazute (teoretice).
Coincidenta se afla in corelatie inversa cu interferenta, si anume: cand coincidenta este egala cu 0, interferenta este absoluta, iar cand coincidenta este de 100% (sau egala cu1,0),interferenta este egala cu 0.
5. ALCǍTUIREA HǍRTILOR CROMOZOMALE
Harta cromozomala constituie reprezentarea grafica a cromozomilor cu indicarea ordinei si distantei relative a genelor linkage (pe fiecare cromozom). Daca distanta dintre gene este estimata prin frecventa crossoverelor (in unitati Morgan), reprezinta o harta genetica. In cazul ca pentru localizarea genelor pe cromozomi se folosesc observatii citologice ale crossoverelor asociate cu mutatii, dislocatii cromozomale etc., se obtine harta citologica. Cele mai complete, sunt hartile citogenetice,care se alcatuiesc pe baza studiilor citologice ale crossoverelor, asociate cu stabilirea genelor de pe fiecare cromozom si a distantelor dintre ele (proportionale cu frecventa crossoverelor).
Alcatuirea hartilor genetice presupune o serie de etape, si anume:
delimitarea grupelor linkage prin identificarea unor gene marker in fiecare cromozom (gene care se transmit in bloc), folosind fenomenul de aneuploidie;
stabilirea distantei dintre genele din acelasi cromozom prin determinarea frecventei crossoverelor;
determinarea ordinei (pozitiei) genelor in cromozomi.
5. 1. DETERMINAREA DISTANTEI DINTRE GENE
Fiecare gena ocupa un anumit locus pe lungimea cromozomului. Pe acelasi cromozom genele sunt dispuse linear, precum margelele intr-un colier. Unitatea de masura cea mai indicata pentru a aprecia distanta dintre doua gene este centimorganul, definit ca fiind partea de cromatida cu probabilitatea de 1% de a suferi o remaniere (crossing over).
Exemplu: Daca genotipul AB // ab da nastere la 8% gameti recombinati Ab si 8% gameti recombinati aB, distanta dintre A si B este de 16 unitati; daca distanta intre B si C este de 12 unitati, inseamna ca 12% din gametii genotipului BC // bc au fost recombinati, adica 6% Bc si 6% bC.
Fiecare crossing over da nastere la doua cromatide recombinate din patru, adica la jumatate.
A) Estimarea distantei intre doi loci
Metoda cea mai simpla pentru decelarea gametilor recombinati la un dublu heterozigot consta in analiza descendentei testcross. Sa presupunem ca in descendenta testcross a unui dublu heterozigot AC // ac au rezultat 37% indivizi cu fenotipul AC, 37% ac, 13% Ac si 13% aC. Indivizii cu fenotipul Ac si aC (genotipurile Ac // ac, si aC // ac) au rezultat din gameti recombinati ai parintelui dublu recesiv. Astfel, 26% (13% + 13%) din gameti sunt recombinati, iar distanta dintre A si C este egala cu 26 unitati.
B) estimarea distantei dintre trei loci
In mod obisnuit, la distante mai mici de 5 unitati intre doua gene, nu se observa crossing overe duble. Pentru decelarea crossing overe-lor duble la distante mai mari este judicioasa folosirea unui marker intre celedoua gene. Sa presupunem ca am facut testcross-ul unui triplu heterozigot ABC // abc si ca descendenta este constituita din:
36 % ABC // abc 9 % aBC // abc 4 % ABc// abc 1 % AbC // abc
36 % abc // abc 9 % Abc // abc 4 % abC// abc 1 % aBc // abc
72% 18% 8% 2%
combinatii crossing over crossing over crossing over dublu
parentale simplu intre simplu intre (1 intre A si B si
A si B B si C 1 intre B si C)
Pentru stabilirea distantei intre A si B se tine cont de toate crossing overele ce au loc intre aceste gene, adica 18% + 2% = 20%. In acelasi mod, distanta B - C este egala cu 8% + 2% = 10%.
Distanta A - C este egala cu 20% (A - B) + 10% (B - C) = 30%. Aceasta distanta ar fi fost de numai 26% daca nu se tinea cont de frecventa crossing overe-lor duble.
5. 2.ORDINEA GENELOR IN CROMOZOM
Aditia (insumarea) distantelor dintre loci permite plasarea genelor in ordinea lor corecta. Daca se considera trei gene inlantuite, exista trei pozitii posibile ale acestora in cromozom, in functie de gena situata intre cele doua. Daca nici un crossing over dublu n-a fost inregistrat, distantele sunt perfect aditive. Cunoscand distantele A - B = 12, B - C = 7 si A - C = 5 se poate determina precis ordinea celor trei gene. Distanta A - b fiind mai mare , ne determina sa apreciem ca gena C este situata intre A si B. Intradevar, A - C (5) + C - B (7) = A - B (12).
A____5____C C__________7____B
A______________12_______________B
Daca am considera ca A sau B se gasesc la mijloc, se observa ca distantele ar fi incompatibile intre ele.
| Contact |- ia legatura cu noi -| | |
| Adauga document |- pune-ti documente online -| | |
| Termeni & conditii de utilizare |- politica de cookies si de confidentialitate -| | |
| Copyright © |- 2024 - Toate drepturile rezervate -| |
|
|
|||
|
|||
|
|||
Proiecte pe aceeasi tema | |||
|
| |||
|
|||
|
|
|||