|






 |  |

| Medicina | Medicina veterinara | Muzica | Psihologie | Retete | Sport |
Medicina
|
|
Qdidactic » sanatate & sport » medicina Etapele raspunsului imun |
Etapele raspunsului imun
ETAPELE RASPUNSULUI IMUN
Elaborarea raspunsului imun este, in esenta, rezultatul cooperarii a doua categorii de celule:
celulele accesorii ale raspunsului imun si celulele limfoide.
R Raspunsul imun se caracterizeaza prin succesiunea urmatoarelor etape:
patrunderea antigenului in organism si inglobarea lui de
catre celulele prezentatoare de antigen;
- prelucrarea antigenului si prezentarea epitopilor pe suprafata celulelor accesorii;
- recunoasterea specifica a componentelor nonself
- activarea limfocitelor si diferentierea celulelor efectoare;
- producerea efectorilor raspunsului imun-faza efectorie
- faza de declin (homeostazie)
- faza de memorie
1.Prelucrarea antigenului si prezentarea epitopilor pe suprafata celulelor accesorii
Prelucrarea si prezentarea antigenelor este o etapa obligatorie a elaborarii raspunsului imun, ce deriva din faptul ca limfocitele T nu interactioneaza direct cu antigenele in stare nativa, ci numai dupa ce acestea au fost prelucrate si prezentate pe suprafata unei celule
- Din punct de vedere biochimic, prelucrarea semnifica:
deplierea, clivarea proteinelor si generarea peptidelor, ca rezultat al unei proteolize partiale.
- Prelucrarea antigenului exogen de catre celulele prezentatoare, parcurge urmatoarele etape:
- internalizarea antigenului in veziculele membranare acide;
- proteoliza partiala;
- cuplarea cu moleculele CMH II;
- transportul la nivelul membranei plasmatice.
- Mecanismele moleculare ale prelucrarii antigenelor sunt putin cunoscute. In macrofage se produce o digestie selectiva a antigenului, in urma careia o parte din epitopi se pastreaza, dar cea mai mare parte a antigenului este complet degradata
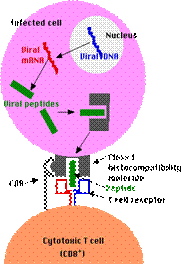
- Prelucrarea antigenului endogen- ex.proteine codificate de gene virale care au infectat celula sau celule infectate viral sunt fagocitate
sunt degradate la peptide in proteazomi,transportate in reticul endoplasmic, cuplare cu CMH I
ANTIGENE ENDOGENE
proteine aberante codificate de gene mutante-cum sunt genele din celulele cancer
proteine codificate de genele virusurilor care au infectat celula -sintetizate in citosol, degradate in proteazomi in peptide virale, transportate in membrana reticul endoplasmic si prezentate limfocitelor Tc. In asociere cu molec MHC clasaI
antigene de pe celulele allogrefelor
Antigene exogene
EXOGENE care intra in corp din mediul inconjurator:
macromolecule inhalate (e.g., proteine -par pisica- declanseaza astm
macromolecule ingerate (proteine-alimente- raspuns alergic
molecule introduse prin piele-injectate ( vaccin) 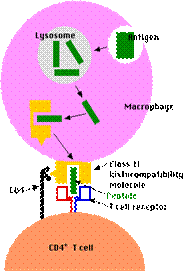
Prelucrarea atg.exogen de catre CPA:
internalizarea atg. in veziculele membr.acide
proteoliza partiala
cuplarea cu molec. MHC II
transportul la nivel membr. plasmat. pt. a putea fi prezentate ly TCD4
Prezentarea epitopilor pe suprafata celulelor accesorii
Celulele prezentatoare de antigen (APC)
constituie o populatie de leucocite cu o capacitate stimulatorie foarte eficienta
sunt gasite in principal in piele, ganglionii limfatici, la nivelul epiteliilor celor mai multe mucoase si in timus.
sunt reprezentate de macrofage, celulele dendritice (interdigitate si foliculare), limfocite B (mai ales cele activate).
Celulele B ca celule prezentatoare de antigen:
cand antigenul se gaseste in concentratii foarte mici,
celulele B cu receptori cu afinitate inalta (IgM sau IgD) sunt cel mai bune CPA deoarece pot captura destul antigen.
celulele B primesc semnalul 1 pentru activare prin legarea antigenului la receptorul BCR,
apoi internalizeaza antigenul si il proceseaza la nivel de peptide mici asociate cu MHC-II,
pe care le prezinta apoi celulelor T helper.
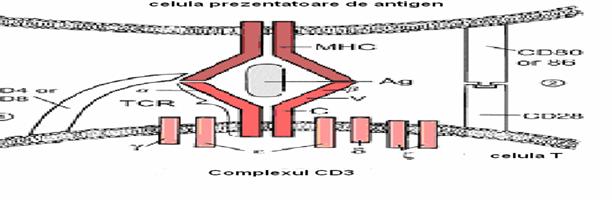
Sa Structurile de pe limfocitele T(TCR si complexul CD3) sunt implicate in procesul de recunoastere a peptidului antigenic.
· Lanturile αβ sau γδ cu domenii constante si variabile(3 regiuni hipervariabile) determina specificitatea pentru legarea asocierii peptid antigenic-CMH.
Lanturile γ δ ε ζ η (polipeptide transmembranare), asociate sub forma de dimeri (complex CD3) au rol in exprimarea TCR si in transductia semnalului antigenic.
2. Recunoasterea specifica a componentelor nonself
TCR si Coreceptorii CD4 si CD8-
recunosc complexul peptid antigenic-MHC de pe APC sau celula tinta
Semnalele biochimice care conduc la activarea ly T sunt declansate de un set de proteine care sunt legate de CD3-lanturile zeta si de coreceptorii CD4 si CD8
Deci functia de receptor al peptidului atgenic este realizata de portiunile V ale lanturilor alfa si beta din TCR, iar semnalizarea este realizata de CD3-lanturi zeta
|
Moleculele de adeziune leaga LyT de APC: ex LFA 1(leucocyte function associated atg )-ICAM1, permite recunoasterea de semnale secundare transmise de APC.
Recunoasterea antigenului este conditionata de existenta fenomenului de histocompatibilitate, adica celulele care prezinta antigenul si cele care il recunosc (limfocitele Tc si Th) trebuie sa poarte molecule CMH identice, adica celulele care interactioneaza trebuie sa apartina aceluiasi organism
Acesta este fenomenul de restrictie (limitare) a interactiunilor celulare prin moleculele CMH
3.Activarea limfocitelor si diferentierea celulelor efectoare
Procesul de activare a ly T implica semnale biochimice generate in urma recunoasterii atg care sunt traduse in raspunsurile biologice ale limfocitelor
In activarea celulelor T intervin molecule(de adeziune) accesorii cu rol in adeziune, recunoastere, semnalizare, de pe celulele T care se leaga la diferiti liganzi de pe APC-denumiti costimulatori .
Integrine-mol de adeziune de pe T naive :
LFA1recunoaste costimulatorul ICAM 1si se elibereaza semnale pt.activarea ly T;
receptorul CD28 de pe LyT recun costimulatorul B7-1 sau B7-2 de pe APC; ac semnale se alatura semnale realizate de TCR si CD4, CD8
Dupa activare pe ly T este exprimat CTLA-4 (CD152)-cytotoxic T lyassociated atg 4, care este un ligand alternativ pentru B7. In momentul legarii CTLA-4 la B7, celula T nu mai primeste semnalul de activare
setul de moleculeCD40L(ly)-CD40(de pe APC)-activarea APC sa exprime mai multe molecule B7 si sa secrete IL-12 care intervin in activarea si diferentierea Ly T
Recunoasterea atg si costimulatorilor de catre LyT initiaza un set de raspunsuri (productia IL-1 de Mf-, secretia de IL-2 cu actiune autocrina -stimuleaza exprimarea RIL-2, proliferarea) care culmineaza cu expansiunea clonelor de limfociteT atg specifice si cu
diferentierea celulelor T naive in celule T efectorii si Tde memorie
stimuleaza activarea si proliferarea limfocitelor B specifice, in functie de natura antigenului.
Mecanisme biochimice de activare a celulei T
Regiunea de contact LyT-APC-sinapsa imunologica:
coreceptorii CD4, CD8 activeaza tirozin protein kinaza atasata la coada citoplasmatica a acestora,
are loc fosforilarea tirozinei
stimularea producerii unor intermediari biochimici care intervin in
activarea factorului nuclear de transcriptie NF-Kb
care stimuleaza transcriptia genelor ce codifica proteinele cu rol mediator in raspunsurile limfocitelor T
Proliferarea si diferentierea celulelor T naive in celule efectorii si de memorie
Celulele Tnaive(Tho)activate secreta IL-2 si exprima IL-2R cu afinitate inalta care leaga IL-2 si initiaza proliferarea celulelor T
Dupa 1-2 zile de la activare are loc expansiunea clonei de limfocite T atg. specifice din care sunt generate limfocitele efectorii TCD4+ si TCD8+.
Dezvoltarea limfocitelor Th1/Th2 nu este un proces intamplator, el este reglat de stimulii pe care ii primesc celulele T naïve cand intalnesc antigenele diferitilor microbi.
Macrofagele si celulele dendritice raspund la multe virusuri si bacterii prin producrea de IL-12 care promoveaza diferentierea celulelor Tho in Th1.
In alte infectii cum sunt cele produse de helminti, celulele T produc ele insasi IL-4 care induce diferentierea spre Th2
In raspunsul fata de stimuli, limfocitele Th antigen specifice, secreta proteine numite citokine cu functii diferite corespunzand celor2 subseturi: Th1(IFN γ,IL2, GM-CSF) sau Th2 (IL3,IL4, IL5, IL6, IL10).
Unele citokine functioneaza impreuna cu antigenul si cu semnalele secundare derivate de la microbi pentru stimularea proliferarii lyT antigen specifice,
altele activeaza CPA care devin competente pentru a stimula expansiunea si diferentierea celulelor TCD8.
Ly TCD8 activate prin antigen si costimulatori se diferentiaza in lyT citolitice(efectorii) care ucid celulele infectate viral, alogrefe, celule neoplazice prin intermediul unor proteine secretate(perforine, granzime).
6. Faza de declin
Odata microorganismul indepartat, mecanismele de amplificare ale raspunsului imun dispar prin feedback negativ
Stingerea raspunsului imun apare in principal ca rezultat a 2 factori
- eliminarea stimulilor pozitivi generati de prezenta microorganismelor
- activarea limfocitelor cu functii
supresoare (limfocite T supresoare).
Dupa stingerea raspunsului imun, - va persista o populatie reziduala de limfocite, celule de memorie, cu specificitate inalta pentru antigenul implicat, care au o durata lunga de viata.
Limfocitele de memorie sunt atat de linie B, cat si de linie T.
Ambele subtipuri celulare asigura:
- protectia oferita de vaccinare,
- raspunsul imun secundar si
- rejectia accelerata a grefei la primitorii de grefe multiple.
7. Faza de memorie
Reactiile din cadrul raspunsului secundar, rapide si explozive, sunt consecinta memoriei imunologice care se instaleaza dupa prima intalnire cu agresorul.
- Stimulul primar antigenic determina proliferarea limfocitelor B si T specifice antigenului; o parte din aceste celule vor evolua spre celule cu functii efectorii iar altele vor deveni celule de memorie care vor interveni in cazul unei noi agresiuni din partea aceluiasi tip de antigen.
- Intensitatea reactiilor de aparare in acest caz va fi mult mai violenta datorita numarului mult mai mare de celule care intra in actiune.
- Limfocitele T si B purtatoare de informatie antigenica conserva memoria: clona care prolifereaza angajeaza un raspuns imun si pastreaza memoria antigenului la nivelul genomului.
- Limfocitele cu memorie recircula fara incetare intre sange, organe limfoide si vase limfatice, capabile de a astepta antigenul strain respectiv, de a-l recunoaste si de a-i raspunde specific.
- Memoria imunologica este independenta de nivelul anticorpilor, este specifica si poate dura ani de zile.
Raspunsul imun umoral
Limfocitele B recunosc atat forma prelucrata cat si forma nativa, nemodificata a antigenului solubil.
Limfocitele B recunosc si leaga specific, polipeptide mici si mijlocii.
Moleculele proteice mari si antigenele corpusculare sunt inglobate si prelucrate de macrofage si de celulele dendritice
Antigenul legat la suprafata limfocitului B, prin intermediul receptorului imunoglobulinic specific, este endocitat, prelucrat in compartimentul acid si prezentat in asociatie cu moleculele CMH II, pentru a fi recunoscut de limfocitele T.
Antigenele T independente declanseaza raspunsul imun prin activarea directa a limfocitelor B, fara sa necesite cooperarea limfocitelor Th. polizaharidul capsular de Streptococcus, ficolul (polimer de sucroza), dextran-sulfatul, LPS ale bacteriilor Gram negative, levanii (polimeri de fructoza), polivinil-pirolidona.
Mecanisme biochimice de activare a celulelor B
Activarea celulelor B cere semnale secundare din care multe sunt produse de-alungul reactiilor de aparare innascute la microbi.
Semnalele initiate de receptorul pentru antigen sunt traduse prin proteinele asociate receptrului (Ig alfa, Ig beta)
Domeniile citoplasmatice ale Ig alfa, Ig beta contin un receptor pentru tirozina .cand 2 sau mai multi receptori BCR sunt grupati , tirozina este fosforilata de catre protein kinazele asociate cu complexul BCR
.Aceste fosfotirozine devin sedii pentru proteine adaptor care ele insasi recruteaza un numar mare de molecule de semnalizare.
Ca urmare, are loc activarea factorului nuclear de transcriptie care stimuleaza transcriptia genelor care codifica produsi proteici implicati in proliferarea si diferentierea limfocitelor B
DINAMICA RASPUNSULUI IMUN
Fazele raspunsului umoral primar
Faza lag-lipsa atc
Log-crestere rapida titru atc
Platou-nivel atc constant
Declin-nivel atc scade prin lipsa sintezei si catabolism
Anticorpii IgM care apar in cadrul raspunsului imun primar (dupa stimulul unic antigenic) sunt anticorpi cu afinitate mica si aviditate inalta. In cadrul raspunsului primar, primii care apar sunt anticorpii din clasa M (IgM) iar dupa 2 - 3 zile apar IgG si IgA care cresc pe masura ce titrul anticorpilor IgM incepe sa scada; IgM dispar dupa cateva saptamani din circulatie.
Raspunsul imun umoral secundar
timp de latenta (inductie) redus - raspuns precoce;
extinderea fazelor in platou si de declin;
rata rapida de sinteza a anticorpilor;
titru inalt al anticorpilor care persista indelung in circulatie;
afinitatea anticorpilor mult mai inalta
CLASIFICAREA ANTICORPILOR ANTIBACTERIENI SISTEMICI:
dupa mecanisme de actiune
activarea fagocitozei bacteriene de catre granulocitele neutrofile, prin opsonizare (anticorpii opsonizanti 'pregatesc' specific suprafata germenului - antigen pentru a putea fi inglobat mai usor in celula fagocitara)
stavilirea efectelor antifagocitare ale unor factori antifagocitari bacterieni (capsula, proteina m streptococica etc. ), prin legare specifica si neutralizare
inhibarea invaziei prin blocarea specifica a factorilor corpusculari si enzimatici de virulenta: capsula, coagulaza, fibrinolizina, lecitinaza, hialuronidaza
neutralizarea exotoxinelor: tetanica, difterica, botulinica, ale clostridiilor gangrenei gazoase, eritrogena streptococica
inhibarea proliferarii dincolo de poarta de intrare, prin activare complement si blocarea transportarii substantelor nutritive
dupa tehnicile de detectare:
- aglutinine: anti-salmonella typhi (reactia widal); anti-rickettsia prowazecki (reactia weil-felix); anti-proteina m streptococica (reactia map); anti-brucella (reactia wright)
- precipitine: anti - antrax; anti-toxina difterica; anti-treponema (vdrl)
- anticorpi neutralizanti: aslo; anti-toxina difterica; anti-toxina tetanica; anti-toxine ale gangrenei gazoase; anti-toxina eritrogena streptococica; anti-toxina botulinica
- anticorpi fixatori de complement: anti-treponema pallidum (reactia bordet-wasermann); anti-brucella; anti-leptospira
- anticorpi detectati prin imunofluorescenta: anti-treponema pallidum
dupa efectele protectoare specifice:
protectori:
- aglutinanti anti-salmonella, anti-rickettsia, anti-proteina M streptococic
neutralizanti anti-toxina eritrogena streptococica, antitoxina tetanica
anti-toxina difterica, anti-toxine ale cl. gangrenei gazoase anti-toxina botulinica
neprotectori:
neutralizanti ASLO, aglutinanti, anti-Brucella
aglutinanti anti-bacil tuberculos
fixatori de complement anti-Brucella si anti-Treponema
precipitanti anti-Treponema
Imunitatea umorala locala
piele si mucoase - imunoglobuline IgA secretorii
MECANISME IMUNE UMORALE SI MEDIATE CELULAR IN RASPUNSUL FATA DE VIRUSURI
RASPUNSUL UMORAL
ANTICORPI IgAs- blocheaza atasarea virusului la celula gazda, prevenind infectia sau reinfectia
IgG, IgM, IgA - bloch. fuziunea anvelopei virale cu membrana plasm. cel.gazda
IgM aglutineaza particulele virale
Complement activat de Ig-mediaza opsonizarea prin C3b si -liza anvelopei virale prin MAC
RASPUNSUL MEDIAT CELULAR
IFN g produs de Th1 si Tc-actiune antivirala directa
Tc distrug celulele self infectate viral
NK, Mf distrugerea celulelor infectate viral, acoperite cu atc prin ADCC
DISFUNCTII IMUNE IN INFECTIA CU HIV
Celulele cele mai sensibile la infectia HIV:sunt celulele care exprima receptori CD4+si coreceptori cum sunt:
ly Th,monocite/macrofage,celule dendritice, microglii cerebrale
Rasp.umoral: activarea policlonala a limfocitelor B prin hiperfunctia Th2-in absenta Th1-se sintetizeaza cantitati crescute de atc neutralizanti,ineficienti,care pot avea chiar efecte deletorii asupra ly CD4+
Rasp. celular -poate controla replicarea virala dupa inf. acuta sau poate ramane la unii bolnavi si in std tarzii ale bolii. In timp are loc depletia LyT.
Depletia celulelor CD4+ (TH1 in special):
efect citopatic al virusului asupra cel CD4+
legarea CD4-gp120, declanseaza apoptoza ly T prin producerea unui semnal inadecvat ptr celula
formarea celulelor giganteT-sincitii multinucleate, nefunctionale,dovedite in vitro,
depltie mediata imunologic -distructia celulelor CD4+(datorita inmugurii virale-liza,=citotoxicitate directa
-lyT neinfectate fix gp120 circulanta pe CD4+al ly T
interventia superantigenelor-proteine retrovirale induc un nr crescut de ly T,care in cazul suprimarii stimularii sunt distruse prin apoptoza
interventia subseturilor Th care normal se controleaza reciproc; HIV perturba interactiunea acestor sisteme conducand la o secretie abundenta de citokine care produce depletia sau anergia ly T
Alterarea exprimarii citokinelor
Diminuarea functiilor celulelor CTL si NK
Cresterea autoanticorpilor si complexelor imune
Diminuarea raspunsului la antigene si mitogene
Alterarea exprimarii MHC-II
Diminuarea chemotaxiei monocitelor
Scaderea intensitatii reactiilor de hipersens. Intarziata (de care raspund Th1)
Continua generare de variante antigenice(prin schimb atg de suprafata)si suprasolicitarea raspunsului imun pot conduce la colaps
Factori implicati in reglarea raspunsului imun
Reglarea raspunsului imun (umoral si celular) este un proces complex de modulare in care intervin o serie de mijloace prin intermediul carora se mentine apararea specifica a organismului la un anumit nivel si cu o anumita durata, in vederea realizarii homeostazei si pastrarii starii de sanatate
Rolul antigenului in reglare
Unele antigene bacteriene activeaza complementul si stimuleaza macrofagele sa exprime molecule co-stimulatorii.
Antigenul stimuleaza r.i. adapativ prin activarea limfocitelor B si T, care la randul lor sintetizeaza anticorpi care activeaza complementul si respectiv citokine care intervin in eliminarea antigenului si care recruteaza alte leucocite
Antigenul este de asemenea un reglator negativ al r.i: cand un atg se leaga la celule imature sau lipsesc semnalele costimulatorii se instaleaza fenomenul de(anergie clonala).
Reglarea prin anticorpi
Anticorpii exercita un control de tip feedback asupra raspunsului imun.
Cand anticorpii circulanti se gasesc in cantitate crescuta competitioneaza cu receptorii membranari pentru antigen, (de natura imunoglobulinica) de pe limfocitele B blocand epitopii si facandu-i inaccesibili receptorilor celulari; ei actioneaza si prin legarea lor de receptorii Fc ai celulelor B, fapt ce inhiba procesul de diferentiere al acestora
Complexele antigen-anticorp circulante aflate in exces raspund de instalarea unor stari imunopatologice grave cum ar fi hipersensibilitatea imediata de tip III; in acest caz raspunsul imun umoral depaseste limitele fiziologice normale.
In anumite conditii conc. scazute de CI se pot lega atat la mIg(ag) cat si la FcR(Atc) ale celulelor B. Aceasta fixare supreseaza productia de anticorpi.
Reteaua idiotip anti-idiotip
Conform teoria retelei imune a lui N. Jerne sistemul imun este prezentat ca o retea idiotip anti-idiotip prin care comunica clonele limfocitare, stimulandu-se sau supresandu-se reciproc. Idiotipul reprezinta un ansamblu de epitopi(idiotopi) lasat de un antigen pe molecula de anticorp corespunzator la nivelul situsului combinativ. Sistemul idiotip-anti-idiotip prezent in ser si pe ly B(BCR) si T(TCR). Anticorpii anti-idiotip vor forma complexe cu idiotipurile circulante corespunzatoare. Cand atc. anti-idiotip se fix de idiotipul BCR conduce la inactivarea celulelor B.
Citokinele Th1 si Th2 isi inhiba reciproc productia si functiile
Echilibrul dintre Th1-Th2 -rol homeostatic asupra raspuns imun
Factori genetici
genele MHC- rol important in recun. atg. de catre cel T
-controleaza rasp. imun fata de agenti infect si fata de atg. proprii modificate. Genele fara relatie cu MHC:- gene care controleaza raspunsul in anticorpi, care regleaza activit macrofage, prolifer si diferentierea ly B etc.
Modificari ale acestor gene influenteaza RI, conducand la imunodeficiente sau la rasspunsuri imune aberante.
Reglare prin retele neuroendocrine(interactiuni sistem imun- SNC-sistem endocrin)
Sist. imun si sist. neuroendocrin comunica prin hormoni si citokine care se leaga de receptorii prezenti in ambele sisteme.
| Contact |- ia legatura cu noi -| | |
| Adauga document |- pune-ti documente online -| | |
| Termeni & conditii de utilizare |- politica de cookies si de confidentialitate -| | |
| Copyright © |- 2025 - Toate drepturile rezervate -| |
|
|
|||
|
|||
|
|||
Lucrari pe aceeasi tema | |||
|
| |||
|
|||
|
|
|||