|






 |  |

| Biologie | Botanica | Chimie | Didactica | Fizica | Geografie |
| Gradinita | Literatura | Matematica |
Biologie
|
|
Qdidactic » didactica & scoala » biologie Analizatorul vizual - globul ocular |
Analizatorul vizual - globul ocular
ANALIZATORUL VIZUAL
Analizatorul vizual receptioneaza, conduce si analizeaza stimulii luminosi de la distanta, permitand organismului sa reactioneze adecvat la cele mai diverse situatii care apar in mediul inconjurator. Impreuna cu analizatorul chinestezic si acusticovestibular, participa la orientarea
organismului in spatiu si la mentinerea echilibrului. Receptioneaza o cantitate impresionanta de informatii din mediul exterior, jucand un rol important in mentinerea si cresterea tonusului activitatii scoartei cerebrale.
Analizatorul vizual are segmentul periferic (retina) situat in ochi Ochiul sau organul vederii este format din globul ocular si din anexele sale (de protectie, de miscare, glandulare).
1. GLOBUL OCULAR
Globul ocular (bulbus oculi) este alcatuit din trei tunici suprapuse concentric, formand peretele globului ocular si din mediile transparente.Cele trei tunici sunt: tunica externa, fibroasa; tunica mijlocie, vasculara si tunica interna, nervoasa, fotosensibila.
1.1. Tunica fibroasa
Tunica fibroasa are doua portiuni: o portiune orala, transparenta - corneea si o portiune aborala, sclerotica, mult mai intinsa, cu rol de sustinere si de protectie. Intre cele doua portiuni se interpune o zona de tranzitie, denumita limb (limbus) sclerocorneean. La animalele de ferma, corneea ocupa 17 - 30 % din suprafata globului ocular, in timp ce la animalele nocturne apare mai intinsa, ajungand pana la 35 %.
Corneea apare ca o lentila convex-concava, complet lipsita de vase sangvine, prezentand variati ale grosimii (0,56 - 1,00 mm) de la o specie la alta. La bovine, canide, felide si suine, corneea apare mai groasa in centru decat la periferie, in timp ce la cabaline este invers.
In structura corneei intra cinci straturi, dintre care doua anterioare, de origine ectodermica (epiteliul anterior si membrana bazala anterioara) si trei posterioare, de origine mezodermica (substanta proprie, membrana bazala si epiteliul posterior).
Epiteliul anterior are 4 - 12 randuri de celule, fiind un epiteliu stratificat pavimentos moale. Desi este expus intermitent contactului cu aerul atmosferic, nu se cheratinizeaza, fiind umezit continuu de pelicula de lacrimi. Celulele epiteliale stabilesc intre ele jonctiuni prin interdigitatii si numerosi desmozomi, formand un strat generator, un strat intermediar si unul superficial. Stratul generator este bogat in glicogen. Celulele epiteliului corneean contin numeroase microfilamente, putin reticul endoplasmic rugos si un complex Golgi redus. Cu cat devin mai superficiale, se aplatizeaza. Celulele stratului superficial prezinta numerosi microvili care au probabil rolul sa retina pelicula de lacrimi pe suprafata corneei. Epiteliul anterior este bogat inervat de numeroaseterminatii libere, care stabilesc contacte cu celulele epiteliale, actionand ca receptori ai durerii.
Capacitatea regenerativa a epiteliului anterior este remarcabila, asigurand o refacere rapida a leziunilor. Exista diferente interspecifice legate de sensibilitatea corneei la injurii si rapiditatea fenomenelor reparatorii, in timpul carora se produce o vascularizare a corneei. La rumegatoare, corneea este putin sensibila la injurii si se cicatrizeaza rapid.Corneea felidelor se cicatrizeaza lent, chiar si dupa o minima scarificare.La canide se instaleaza o neovascularizare intensa, urmata de o cicatrizare rapida. Corneea de la cabaline este cea mai sensibila, reactioneaza sever la impulsuri si se cicatrizeaza lent.
Membrana bazala anterioara este formata din lamina densa si lamina reticularis, ce cuprinde o retea neregulata de fibre reticulare. Dupa Dellmann si Collier- 1987, membrana bazala anterioara nu trebuie confundata cu lama limitata anterioara (membrana Bowmann), ce
reprezinta o modificare a stratului superficial al substantei proprii.
Substanta proprie (substantia propria), stroma sau "dermul" corneei este un tesut conjunctiv fibros lamelar, ce cuprinde circa 100 de lamele (la felide) sau straturi de fibre de colagen, paralele cu suprafata corneei. In interiorul unei lamele, fibrele de colagen sunt paralele intre ele, dar in lamelele invecinate au directii diferite. Legatura dintre fibrele unei
lamele si dintre lamele se realizeaza printr-o substanta fundamentala redusa cantitativ, ce contine glicozaminoglicani sulfatati (condroitin sulfat, keratan sulfat, acid hialuronic). Aceasta substanta asigura transparenta corneei prin mentinerea unui grad optim de hidratare, cunoscut
fiind ca o crestere a cantitatii de apa produce opacifierea corneei. Totodata, impiedica vascularizarea corneei in conditii normale.
Intre lamele sunt prezente fibroblaste, cu aspect stelat, cu putina citoplasma si putine prelungiri ce vin in contact unele cu altele. Spre limbul sclerocorneean sunt prezente si histiocite, iar numai ocazional fibre elastice.
Membrana bazala posterioara apare ca o structura subtire, PAS-pozitiva, mai retractila si mai fragila decat membrana anterioara. Ultrastructural, apare formata din fibrile de colagen, lipsite de periodicitatea caracteristica, dispuse in arii hexagonale regulate. Membranele bazale
se pot regenera pe seama epiteliilor (anterior si posterior).
Sclerotica. In structura scleroticii se disting trei straturi: episclera,substata proprie si lamina fusca. Episclera este stratul exterior, format din tesut conjunctiv elastic - lax, cu numeroase vase de sange. Substanta (tesutul) propriu este format din tesut conjunctiv dens, sarac in elemente elastice si vase proprii. Fasciculele de fibre elastice sunt dispuse longitudinal, paralel cu suprafata globului, dar se intretes strans si se dispun circular la nivelul limbului sclerocorneean si in jurul nervului optic. Lamina fusca este stratul intern, de tranzitie spre coroida, in care predomina fibrele elastice, fibroblastele si melanocitele.
Pe sclerotica se insera muschii oculomotori ale caror fibre tendinoase se intretes cu fibrele sclerale. Sarac vascularizata, sclerotica este perforata de vase de tranzit, fiind aparent lipsita de capilare limfatice, dar prezentand spatii limfatice. Este inervata de fibre mielinice si amielinice, ce se termina in organe asemanatoare cu fusurile neurotendinoase.In grosimea scleroticii s-au semnalat si celule ganglionare. La batracieni,sclerotica eate formata din tesut cartilaginos, la pasari din tesut cartilaginos hialin care se poate osifica la adulte, iar la reptile din tesut osos.
La pasari, in apropierea corneei, se structureaza un inel cartilaginos cranial (care se osifica la speciile rapitoare, inel care preia functia de sprijin si protectie a orbitei). Un inel scleral caudal, cartilaginos, inconjoara total sau partial nervul optic.
Limbul sclerocorneean (limbus) reprezinta zona de trecere dintre cornee si sclerotica. Epiteliul anterior ai corneei se continua cu epiteliul conjunctival, care se sprijina pe un tesut conjunctiv lax. Substanta proprie a corneei se continua cu substanta proprie a scleroticii. Lamele de fibre colagene se asociaza cu fibre elastice si se continua cu fibrele ecuatoriale ale scleroticii. La nivelul contactului cu marginea ciliara a irisului, membrana bazala posterioara se desface intr-un sistem de lamele care se anastomozeaza cu fibrele conjunctivo-elastice din stroma irisului,formand ligamentul pectinat (ligamentum pectinatum) al irisului, foarte dezvoltat la bovine si cabaline. Intre fibrele ligamentului pectinat se delimiteaza o serie de spatii intercamunicante (Fontana), ce se deschid printr-o serie de orificii in camera anterioara. Spatiile intercomunicante sunt captusite cu un endoteliu, aflat in continuitate cu endoteliul camerei anterioare si al sinusulu si venos scleral. In grosimea limbului se gaseste o bogata retea vasculara ce formeaza un inel vascular prericorneean, care asigura prin difuziune nutritia corneei.
Canalul Schlemm, prezent la om, este un spatiu triunghiular neregulat, tapetat de endoteliu, situat in marginea posterioara a limbului, adiacent unghiului iridocorneean (spatium anguli iridocornealis). Intre canal si camera anterioara se interpune o retea de fine trabecule conjunctive (reticulum trabeculare), tapetate cu celule endoteliale. Umoarea apoasa este "filtrata" prin aceste spatii intertrabeculare inainte de a ajunge in canalul Schlemm. Nu exista comunicatii directe intre spatiile trabeculare, incat resorbtia umoarei apoase se realizeaza prin dializa, umoarea trecand prin cele doua straturi endoteliale si tesutul conjunctiv dintre ele. Perturbarea resorbtiei produce glaucomul, in urma cresterii presiunii intraoculare. Canalul Schlemm dreneaza umoarea apoasa prin canale in sistemul venos episcleral, datorita unui gradient de presiune ce impiedica refluxul in canal. Acest canal nu exista la animalele domestice.
1.2. Tunica mijlocie
Tunica mijlocie a globului ocular, tunica vasculara (tunica vasculosa bulbi) sau tractusul uveal (uvea) are trei portiuni: coroida, corpii ciliari si irisul.
Coroida (choroidea) reprezinta segmentul aboral al tractusului uveal, aparand ca o membrana conjunctivo-vasculasa si pigmentara,interpusa intre sclerotica si retina vizuala. In structura coroidei se disting patru straturi: epicoroida, stratul vascular, stratul coriocapilar si
membrana bazala.
Epicoroida sau supracoroida (lamina suprachoroidea) este stratul cel mai extern, care se confunda practic cu lamina fusca a scleroticii. Epicoroida este formata din fibre de colagen si elastice dispuse in lamele reticulare. Fascicule de fibre de colagen se desprind indreptandu-se spre sclerotica si continuandu-se in aceasta. Intre aceste fibre se delimiteaza numeroase spatii pericoroidale, care impreuna cu lacunele intercomunicante ale epicoroidei sunt umplute cu lichid cerebro-spinal. Populatia celulara a coroidei cuprinde fibroblaste, numeroase melanocite si ocazional macrofage.
Stratul vascular cuprinde numeroase artere si vene. Venele au pereti subtiri si sunt foarte numeroase, cele mai mari fiind situate extern, iar cele mijlocii, intern. Venele au traiect puternic spiralat, fiind vorticoase. Arterele sunt mai putin numeroase, mai mici, situate profund si au un perete muscular evident. Tesutul conjunctiv din stratul vascular este format din lamele conjunctive, fibre elastice, rare leiocite si numeroase melanocite, ramificate si anastomozate.
Stratul coriocapilar (lamina chroidocapilaris) este format dintr-o retea de capilare largi (cu lumenul intre 10 - 15 mm), dispusa intr-un singur plan. Asigura nutritia coroidei. In jurul regiunii maculare, capilarele sunt foarte largi si realizeaza numeroase anastomoze venoase. Printre capilare se gasesc fibre de colagen, elastice, si numeroase celule conjunctive .
Capilarele prezinta un epiteliu fenestrat, iar nucleii celulelor endoteliale si ai pericitelor sunt dispersati numai pe fata externa a stratului,incat peretele capilar fuzioneaza cu membrana bazala a epiteliului pigmentar al retinei, favorizand schimburile cu aceasta.
Complexul bazal (complexus basalis) sau membrana bazala (Briich) delimiteaza coroida de retina. Variaza ca dezvoltare si grosime de la o specie la alta. Atunci cand este complet dezvoltat, complexul bazal cuprinde cinci structuri: membrana bazala a epiteliului pigmentar al retinei; o lama interna de natura colagena; un strat elastic; o lama colagena externa; membrana bazala a stratului corio-capilar. In regiunea tapetului lucid, complexul bazal este format numai din fuzionarea celor doua membrane bazale, lipsind celelalte straturi.
Tapetul lucid (tapetum lucidum), prezent la carnivore, rumegatoare si ecvine, este un strat zonal al coroidei, care reflecta lumina, permitand o imbunatatire a perceptiei optice in lumina slaba. Ocupa jumatatea dorso-laterala a fundului de ochi, fiind dispus intre stratul coriocapilar si stratul vascular al coroidei. Lipseste la hominide, suine si pasari.
Forma, marimea, culoarea, intinderea si structura variaza de la o specie la alta.
- La ierbivore (rumegatoare, ecvine), tapetul este fibros (tapetum fibrosum), fiind constituit dintr-o intretaiere de fibre conjunctive, irizate, cu traiect sinuos si putine fibroblaste, leiocite si melanocite. Fibrele de colagen sunt intretesute dens, fiind dispuse paralel cu suprafata retinei.
- La carnivore, tapetul este celular (tapetum cellulosum), fiind format din mai multe straturi de fibroblaste poligonale, aplatizate, incarcate cu o mare cantitate de zinc. La feline, celulele tapetale contin si o mare cantitate de riboflavine. Numarul de straturi de celule tapetale
este mai mare in zona centrala, ajungand la 15 straturi la canide si 35 de straturi la felide, descrescand la un singur strat la periferie. Celulele fiecarui strat sunt dispuse in tandem, iar axul lor longitudinal este paralel cu retina. Straturile succesive ale tapetului apar delimitate intre ele prin fibre de colagen, elastice si capilare. In citoplasma celulelor tapetului se intalnesc numerosi melanozomi modificati, cu aspect fibrilar,paraleli cu suprafata retinei. Melanozomii contin mult zinc, care contribuie la reflectia luminii. Reflectia luminii se datoreste orientarii spatiale a melanozomilor sau a fibrelor colagene la ierbivore. Stratul epitelial al retinei care acopera tapetul este nepigmentat, fapt ce permite observarea relativa usoara a tapetului.
- La pasari, coroida este foarte pigmentata, iar tapetul lucid lipseste. De pe coroida, la nivelul papilei, se detaseaza spre camera interioara o vilozitate corio-capilara, ce contine celule pigmentare si nevroglii, denumite pieptene (pecten), cu aspect de pana, prezentand o lama
centrala si pliuri secundare. Pectenul apare mai dezvoltat la pasarile rapitoare diurne si la gasca.
Arterele coroidei se ramifica intens, realizand reteaua coriocapilara,din care se desprind venele.
Inervatia coroidei cuprinde filete nervoase vasomotorii, care formeaza plexuri in epicoroida si stratul vascular. Se pot intalni si neuronii ganglionari, implicati in inervatia vegetativa a vaselor.
Rolul coroidei consta in asigurarea nutritiei retinei, in mentinerea si reglarea presiunii intraoculare, in realizarea camerei obscure a globului ocular.
Corpul ciliar (corpus ciliare) reprezinta continuarea in sens oral a coroidei, fiind situat intre coroida si iris. Pe o sectiune longitudinala prin globul ocular, corpul ciliar apare triunghiular, cu baza orientata spre camera anterioara si iris, iar cu varful triunghiului spre coroida. Organizarea morfologica a corpului ciliar reprezinta o adaptare
|
functionala a coroidei in sensul ca muschiul ciliar inlocuieste epicoroida, lipseste stratul corio-capilar si apar procesele ciliare. Ca atare, corpul ciliar este format din muschiul ciliar si procesele ciliare.
Muschiul ciliar (musculus ciliaris) este constituit din fibre musculare netede, dispuse in trei grupe diferit orientate: fibre meridionale, fibre radiale si fibre circulare. La pasari este format din fibre musculare striate
- Fibrele meridionale sau longitudinale, cele mai externe, se intind de la cornee spre coroida, inserandu-se prin tendoane pe complexul bazal al coroidei si actionand ca tensor al coroidei. La suine, fibrele meridionale predomina in zona temporala a globului ocular. La alte specii de mamifere formeaza aproape in intregime portiunea posterioara a muschiului ciliar. La carnivore, practic tot corpul ciliar este format din fibre rneridionale.
-Fibrele radiale (fibrae radiales) se desprind de pe radacina irisului, dispersandu-se in evantai spre fata interna a corpului ciliar. Fibrele circulare sunt mai putin numeroase. Predomina in zona nazala a corpului ciliar, unde la suine sunt singurele fibre existente.
In general, la animalele domestice, muschii ciliari nu sunt bine dezvoltati, incat posibilitatile de acomodare ale cristalinului sunt relativ reduse.
Muschiul ciliar este inervat de fibre vegetative provenite din ganglionul ciliar.
Procesele ciliare sunt structuri secretorii, cu aspect de cute meridionale, care se intind de la nivelul orei serrata pana in apropierea bazei irisului, unde se termina brusc. Sunt formate din ghemuri vasculare venoase si capilare mari, cuprinse intr-o stroma conjunctiva bogata in fibre elastice. Procesele ciliare sunt tapetate de retina oarba, reprezentata de un epiteliu bistratificat, dispus pe o membrana bazala aflata in continuarea complexului bazal al coroidei. Epiteliul proceselor ciliare prezinta un strat epitelial extern, pigmentat, aflat in prelungirea epiteliului pigmentar al retinei, si un strat epitelial intern, nepigmentat.
Stratul epitelial extern, pigmentat, este constituit din celule cuboidale intens pigmentate, care prezinta invaginatii ale plasmalemei bazale. Suprafata opusa polului bazal stabileste interdigitatii si complexe jonctionale cu celulele stratului intern, nepigmentat.
Stratul epitelial intern, nepigmentat, este format din celule cubice sau prismatice, sprijinite pe o membrana bazala ce continua membrana limitanta interna a retinei. Polul bazal al celulelor din acest strat, orientat spre camera posterioara, prezinta numeroase invaginatii plasmalemale profunde, asociate cu mitocondrii. In citoplasma este prezent un reticul endoplasmic rugos bine dezvoltat si un complex Golgi evident. Celulele din acest strat stabilesc jonctiuni gap atat intre ele, cat si cu polul apical al celulelor din stratul pigmentar. Umoarea apoasa reprezinta un produs de filtrare a plasmei sangvine din capilare, produs care este transportat transepitelial in camera posterioara cu ajutorul jonctiunilor gap. Celulele epiteliului nepigmentat dau nastere unor fibre zonulare care se insera pe membrana bazala.
La cabaline si rumegatoare, procesele ciliare sunt lungi, incat devin vizibile la marginea dorsala a pupilei formand "funinginea".
Irisul (iris) reprezinta segmentul cel mai anterior al tractusului uveal, avand aspectul unei diafragme circulare, ce delimiteaza un orificiu central, pupila. Prin pozitia sa, irisul imparte compartimentul anterior al globului ocular in doua camere, anterioara si posterioara, care
comunica intre ele prin pupila.
Irisul prezinta o structura complexa, fiind format din cinci straturi, dintre care primele doua straturi reprezinta o continuare a coroidei, in timp ce ultimele doua, posterioare, reprezinta o continuare a retinei oarbe, ce acopera corpul ciliar. Stratul mijlociu are origine dubla, stroma provenind din coroida, iar sfincterul peripupilar din elementele retinei. Cele cinci straturi sunt: epiteliul anterior, stratul avascular, stratul vascular, stratul muscular si epiteliul posterior.
Epiteliul anterior reprezinta prelungirea endoteliului posterior al corneei, aparand simplu, pavimentos, discontinuu la adult, dar continuu la foetus si nou-nascut. Este format din fibroblaste mari, poligonale si melanocite. Prin discontinuitatile epiteliului anterior se stabilesc comunicari intre camera anterioara si spatiile lacunare din iris, iar la nivelul marginii pupilare, se patrunde in cripte ce ajung in stroma.
Stratul limitant anterior reprezinta stratul avascular al stromei irisului, asemanandu-se cu structura laminei fusca. Este format din fibre colagene si un numar mare de celule pigmentare, fiind totodata bogat in proteoglicani. Acest strat conduce la determinarea culorii ochilor. Cu-
loarea ochilor este dependenta de grosimea stratului limitant anterior, de numarul celulelor melanofore, de gradul lor de pigmentare si de epiteliul posterior pigmentat. Atunci cand stratul limitant este subtire si numarul melanoforelor este mic, lumina este reflectata de melanina din
epiteliul posterior, obtinandu-se o culoare albastra. O stroma mai densa cu un mare numar de celule pigmentare, confera irisului o culoare cenusie. Culoarea neagra sau bruna este determinata de un strat limitant antetior gros si de un numar mare de melanocite. Pigmentatia irisului apare caracteristica la diferite specii de animale domestice. La canide domina culoarea bruna, cu doua tonuri, realizandu-se o heterocromie iriana. La cabaline, irisul prezinta o singura culoare sau o mixtura de culori, cu zonari pigmentare. La ungulate, irisul se caracterizeaza printr-o distribuire relativ uniforma a pigmentului. Hiperpigmentarile irisului pot sugera inflamatii cronice, hipopigmentarea aparand la animalele albinotice.
Stratul mijlociu, stratul vascular sau stroma irisului, intens pigmentat si vascularizat, contine fibre de colagen si elastice, dispuse circular, radiar sau in arcade. In acest strat sunt cuprinse arterele iriene, care provin dintr-un cerc arterial major, dispus la periferie. Un mic cerc vascular peripupilar cuprinde o vena, rezultata din confluarea capilarelor emise de arterele iriene. In iris nu exista vase limfatice, ci numai lacune ce contin limfa.
Tot in grosimea stromei se gasesc fibre musculare netede circulare, care formeaza sfincterul irisului, inervat de parasimpatic. La pasari, acest muschi este striat. Stroma iriana mai contine fibroblaste, mastocite,histiocite si numeroase melanocite. La pesti, celulele conjunctive ale stromei prezinta o structura fibrilara sau contin criste aciculare de guanina,care confera irisului un luciu metalic caracteristic.
Stratul muschiului dilator al pupilei este format din celule mioepiteliale provenite din modificarea unor elemente celulare din retina. Fibrele musculare din iris se intersecteaza unele cu altele in unghiuri ascutite cu diferite valori, pe marginea laterala si mediala a pupilei, la animalele cu pupila ovala sau pe marginea dorsala si ventrala la animalele cu fanta pupilara orientata vertical. Fibrele musculare din iris primesc o dubla inervatie, orto si parasimpatica.In acest fel, irisul actioneaza ca un diafragm care regleaza cantitatea de lumina ce ajunge pe retina si participa la corectia aberantei de sfericitate.
Epiteliul posterior, bistratificat, continua epiteliul corpului ciliar si pe cel pigmentar al retinei, prezentand un strat anterior (sau extern) si un strat posterior (sau intern). Stratul anterior continua stratul corespondent din epiteliul corpului ciliar si participa la formarea muschiului dilatator al pupilei, prin modificarea celulelor componente. Astfel, portiunea bazala a celulelor epiteliale se alungeste, se ramifica si se cuteaza, capatand caracteristici ale miocitelor. Portiunea apicala isi pastreaza aspectele structurale tipice ale celulelor epiteliului pigmentar, de unde denumirea de miopigmentocite. Muschiul dilatator al pupilei este inervat
de ortosimpatic.
Nervii irisului sunt in majoritate fibre para- si ortosimpatice, ce actioneaza asupra fibrelor musculare din iris. Exista, de asemenea, fibre senzitive pe fata anterioara si fibre vasomotoare.
Unghiul iridian sau iridocorneean (angulus iridocornealis) corespunde periferiei camerei anterioare, fiind delimitat intre fata posterioara a corneei si fata anterioara a irisului. La nivelul lui, endoteliul corneean se continua cu epiteliul anterior al irisului. Unghiul iridian prezinta o importanta functionala deosebita pentru circulatia si absorbtia umoarei apoase. La nivelul unghiului iridian exista o retea complicata de trabecule ce delimiteaza intre ele spatiile unghiului (Fontana). Trabeculele sunt formate din fibre de colagen, fibroblaste si melanocite, fiind invelite de celule endoteliale. Inspre camera anterioara, trabeculele formeaza ligamentul pectinat care ataseaza irisul la circumferinta interna a corneei. Prin spatiile Fontana circula umoarea apoasa care este preluata din camera anterioara si drenata prin vene colectoare in plexul venos scleral, la animalele domestice neexistand sinus venos scleral (canal Schlemm).
1.3. Tunica interna
Tunica interna (tunica sensoria bulbi) sau retina are origine nervoasa (diencefalica), prezentand o portiune vizuala, fotosensibila, situata posterior si ocupand 5/6 din circumferinta tunicii, si o portiune oarba, lipsita de elemente fotosensibile, ce tapeteaza corpul ciliar si irisul. Zona de trecere dintre aceste doua segmente se numeste ora serrata.
In timpul dezvoltarii embrionare a veziculelor optice se produce o invaginare a acestora, astfel incat in fiecare vezicula se formeaza doua straturi: un strat intern din care se vor diferentia elementele nervoase ale retinei si un strat extern, care va genera epiteliul pigmentar al retinei ce va ajunge in contact intim cu coroida.
Retina vizuala sau optica (pars optica retinae). Are o grosime care se micsoreaza dinapoi inainte de la 350 µm in vecinatatea papilei optice, la 150 µm in apropiere de ora serrata. Este formata din zece straturi de celule, din care unul singur, epiteliul pigmentar al retinei, este denatura epiteliala, iar celelalte noua sunt de natura nervoasa, fiind constituite din neuroni si nevroglii, ca si din prelungirile acestora.
In structura retinei participa trei tipuri de celule nervoase: celule fotoreceptoare, neuroni bipolari si neuronii multipolari, conectati in tandem. Cele trei tipuri celulare formeaza patru din straturile retinei, datorita faptului ca portiuni ale elementelor vizuale sunt prezente in doua
straturi. Conexiunile sinaptice dintre cele trei tipuri de neuroni formeaza 2 straturi, iar al treilea strat este alcatuit din axonii neuronilor multipolari care formeaza fibrele nervilor optici. Prelungirile celulelor nevroglice structureaza doua membrane limitante, una interna si alta externa.
Celulele pigmentare (pigmentocytus) sunt situate pe complexul bazal al coroidei, ce apare ondulat, luand contact strans cu stratul coriocapilar. Au o forma poliedrica turtita, aparand mai lungi decat inalte. De pe fata lor interna se detaseaza numeroase prelungiri care se interpun intre conurile si bastonasele celulelor vizuale, ajungand pana aproape de membrana limitanta externa. Nucleii, adesea dubli, mari, ovoizi si nucleolati, sunt situati central. Fata bazala a celulelor prezinta invaginari profunde ale membranei plasmatice, intre care apar numeroase mitocondrii. Lateral si paraapical se realizeaza jonctiuni de tipul zonulelor
ocludens si adherens. Citoplasma celulelor granulare contine numeroase granule de fucsina si un reticul endoplasmic (neted si rugos) dezvoltat. In conditii de intuneric, granulele sunt concentrate perinuclear, pentru ca in lumina sa se disperseze in toata citoplasma, inclusiv in prelungiri.
Fucsina favorizeaza concentrarea excitatiilor luminoase la nivelul celulelor vizuale, participand si la formarea rodopsinei. Granulele pigmentare lipsesc in epiteliul care acopera tapetul lucid. Intre celulele pigmentare si prelungirile in forma de con sau bastonas se stabilesc raporturi stranse,in sensul ca fiecare con sau bastonas este cuprins intr-o minuscula camera "obscura", realizandu-se schimburi intre cele doua tipuri de celule (fig. 16.9.).
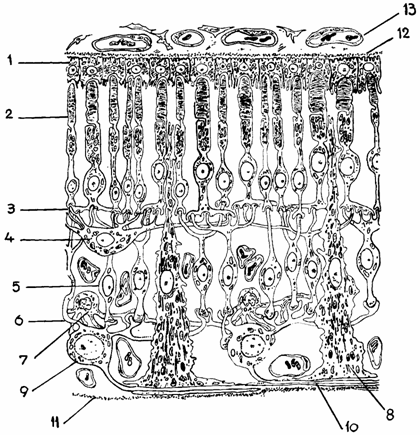
Fig. 16.9. Structura retinei- schema
1 - epiteliul pigmentar; 2 - celule vizuale; 3 - strat plexiform extern; 4 - neuroni orizontali; 5 - neuroni bipolari; 6 - strat plexiform intern; 7 - neuroni amacrini; 8 - nevroglii radiale; 9 - neuroni multipolari; 10 - fibre optice; 11 - membrana limitanta interna; 12 - membrana bazala; 13 - strat coriocapilar.
Celulele vizuale sau fotoreceptoare (epitheliocytus neurosensorius) sunt neuroni modificati morfologic si functional. Au o forma alungita, prezentand: un corp care contine nucleul, o prelungire externa, cu semnificatie de dendrita, in forma de bastonas sau con, si o prelungire interna,relativ rectilinie, cu semificatie de axon. Corpul si nucleul celulelor vizuale se gasesc in stratul nuclear extern. Corpul celular, impreuna cu prelungirea interna, forteaza segmentul intern al celulei fotoreceptoare, in timp ce prelungirea externa reprezinta segmentul extern. Conurile si bastonasele apar lungi la cabaline, dar scurte la rumegatoare si suine.
Cele doua segmente, intern si extern, ale celulelor vizuale sunt situate in straturi diferite ale retinei, fiind despartite intre ele prin membrana limitanta externa.
-Segmentul extern, in forma de bastonas sau con, este alcatuit din doua portiuni sau articole, legate intre ele printr-o piesa intermediara cu structura de cil. Articolul extern reprezinta structura receptoare, intrucat contine pigmentul vizual, o substanta fotosensibila care, absorbind lumina, sufera modificari chimice, generand un potential electric, influxul nervos. Articolul extern sufera un proces continuu de dezintegrare si refacere. Structurile degradate sunt fagocitate de celulele din epiteliul pigmentar, iar refacerea se realizeaza prin procesele de biosinteza, care au loc in articolul intern.
- Segmentul intern al celulelor vizuale cuprinde corpul celulei si axonul, care stabileste contacte sinaptice cu neuronii bipolari. Nucleii celulelor cu bastonase sunt mici si situati la oarecare distanta de membrana limitanta externa, in timp ce nucleii celulelor cu conuri sunt mari,localizati in vecinatatea imediata a membranei.
Celulele vizuale cu bastonase (epitheliocytus neurosensorius bacillifer) isi datoreaza numele, formei de "bastonas" pe care o prezinta segmentul extern. Celulele cu bastonase sunt cele mai numeroase, ajungand la aproximativ 130 milioane, la om.
- Articolul extern al bastonasului prezinta o ultrastructura caracteristica, fiind format din sute de discuri membranoase, rezultate prin plicaturarea plasmalemei. Discurile sunt situate unele peste altele sicontin molecule de rodopsina, care reprezinta pigmentul vizual. Rodopsina se descompune sub actiunea luminii in retinen si o proteina incolora. La intuneric, rodopsina se regenereaza. Daca actiunea luminii a fost de lunga durata, refacerea rodopsinei poate avea loc numai in prezenta vitaminei A. Cantitatea de rodopsina este mult mai mare la animalele cu vedere nocturna, lipsind complet la unele specii de pasari.
- Piesa intermediara, ingustata, uneste cele doua articole ale segmentului extern si are structura unui cil modificat, lipsit de perechea centrala de microtubuli.
- Articolul intern al bastonasului, ovoidal ca forma, prezinta o regiune externa cu striatiuni longitudinale, determinate de prezenta mitocondriilor, si o regiune interna, cu aspect granular, care contine complexul Golgi si reticul endoplasmic (rugos si neted). In articolul intern
au loc procese de biosinteza proteica, asigurand refacerea articolului extern.
Celulele vizuale cu bastonase sunt adaptate pentru vederea in lumina redusa. Ele au un prag foarte mic de stimulare luminoasa, fiind excitate de cea mai mica raza de lumina si inhibate de lumina puternica. Totodata, celulele cu bastonase sunt adaptate pentru diferentierea intensitatilor luminoase. La animalele cu activitate predominant nocturna (soarece, sobolan, bufnita, liliac), retina este formata aproape in exclusivitate din celule vizuale cu bastonas. In fiecare zi, lumina provoaca distrugerea discurilor mai vechi de la extremitatea distala a bastonasului, concomitent cu adaugarea de noi discuri la extremitatea proximala.
Celulele vizuale cu conuri (epitheliocytus neurosensorius conifer) au prelungirea dendritica in forma de con. Apar mai putin numeroase (atingand 6 - 7 milioane la om), fiind singurele prezente in regiunea petei galbene. Articolul extern este mult mai scurt (10 mm) decat celintern (25 mm). Ultrastructural, articolul extern este format din discuri membranoase, care se dezintegreaza si se refac continuu. Pigmentul vizual este reprezentat de iodopsina. Piesa intermediara are structura de cil modificat. Articolul intern al segmentului extern are forma de
bulb, prezentand un aspect striat in regiunea externa, datorita mitocondriilor, si un aspect granular in regiunea interna, contractila, denumita corp mioid. Corpul mioid contine complexul Golgi si reticul endoplasmic rugos si neted, fiind dotat cu proprietati contractile, incat conurile prezinta fenomene retinomotoare scurtandu-se la lumina si alungindu-se la intuneric.La batracieni, corpul mioid contine un corpuscul denumit paraboloid. Totodata, la nivelul articolului intern se realizeaza sinteza de proteine, care asigura refacerea articolului extern.
Segmentul intern al celulei cu con cuprinde corpul celulelor si axonul. Corpul celulei prezinta un nucleu mare, oval, situat in imediata vecinatate a membranei limitante externe. Axonul se indreapta spre stratul plexiform extern.
Celulele vizuale cu conuri au aparut relativ tardiv pe scara filogenetica. Ele se caracterizeaza printr-un ridicat prag de excitabilitate,fiind adaptate pentru vederea diurna si pentru perceperea formelor si culorilor. La reptile nu exista decat celule cu conuri.
Neuronii bipolari (neuronum bipolare), de forma ovoidala, prezinta doua prelungiri: o prelungire dendritica, externa, si o prelungire axonica, interna. Prelungirea dendritica face sinapsa cu axonul unei celule cu con sau cu axonii mai multor celule cu bastonas. Prelungirea axonica, subtire si lunga, stabileste sinapse atat cu neuronii multipolari,cat si cu neuronii amacrini.
Neuronii multipolari sunt neuroni mari, cu nuclei eucromatici, veziculosi. Prezinta mai multe prelungiri dendritice care se ramifica in stratul plexiform intern, stabilind sinapse cu axonii neuronilor bipolari. Axonii neuronilor multipolari trec in stratul fibrelor optice, de unde
se indreapta spre papila nervului optic.
Neuronii de asociatie din retina sunt de doua tipuri: neuroni amacrini si neuroni orizontali.
Neuronii amacrini (neuronum amacrinum) sunt lipsiti de axon, prezentand unul sau doua trunchiuri dendritice scurte, care se ramifica intens, stabilind sinapse cu dendritele neuronilor multipolari si cu axonii neuronilor bipolari. Nucleii neuronilor amacrini prezinta o forma neregulata, cu invaginari profunde ale membranei nucleare.
Neuronii orizontali (neuronum horizontale) sunt celule mari, poliedrice, cu prelungiri dendritice si axonale, dispuse paralel cu suprafata retinei. Dendritele par scurte si stabilesc sinapse cu axonii celulelor vizuale si cu dendritele celulelor bipolare. Axonul lor, lung si subtire, realizeaza sinapse cu prelungirile celulelor vizuale si cu dendritele neuronilor bipolari. Neuronii orizontali sunt dispusi in stratul granular intern.
Celulele de sustinere sunt reprezentate de macroglii si de gliocite radiale. Macrogliile sunt dispuse in stratul fibrelor optice. Gliocitele radiare (gliocytus radialis) sau celulele Müller, sunt elemente nevroglice mari, alungite, cu aspect de fibre, intinse de la membrana limitanta externa pana la membrana limitanta interna. Corpul gliocitelor radiale, situat in stratul nuclear intern, este format dintr-un nucleu, inconjurat de un strat subtire de citoplasma granulara, intens colorata. De la corpul celular se desprind doua prelungiri radiale (processus radialis) una externa si alta interna. De pe aceste prelungiri se detaseaza expansiuni care patrund orizontal in straturile retinei. Aceste expansiuni sunt lameliforme in straturile nucleare (extern si intern) si in stratul ganglionar, formand cosulete in jurul celulelor vizuale si nervoase. In straturile plexiforme (intern si extern) prelungirile sunt filiforme. Nevrogliile radiale Müller ofera suport mecanic si nutritional neuronilor din retina. Ele mediaza transferul unor metaboliti esentiali (exemplu glucoza) catre neuronii retinali.
Diferitele tipuri de celule din retina, impreuna cu prelungirile lor, participa la realizarea celor zece straturi ale retinei.
Stratul pigmentar (stratum pigmentosum) adiacent coroidei, este un epiteliu simplu, cuboidal sau scuamos, format din celule pigmentare. Acest strat indeplineste functii complexe precum: transportul metabolitilor de la sange la celulele vizuale; fagocitarea; degradarea lizozomala si reciclarea segmentelor externe ale celulelor vizuale, absorbitia luminii si crearea unor conditii de obscuritate in camera interioara a globului ocular.
Stratul conurilor si bastonaselor (stratum nervosum) cuprinde segmentele externe (conurile si bastonasele) ale celulelor vizuale.
Membrana limitanta externa (stratum limitans externum), format din extremitatile prelungirilor externe ale glicocitelor radiale (Müller) delimiteaza stratul conurilor si bastonaselor de stratul nuclear extern. Prelungirile radiale ale celulelor Müller prezinta microvili ce proemina intre segmentele externe ale celulelor cu conuri si bastonase.
Stratul nuclear extern (stratum nucleare externum) cuprinde corpul celulelor vizuale, cu nuclei mari, ovoizi, ai celulelor cu conuri, dispusi pe un singur rand in vecinatatea imediata a
membranei limitante externe si cu nuclei mici si rotunzi ai celulelor cu bastonase, dispusi pe 2 - 5 randuri situate in profunzimea stratului.
Stratul plexiform extern (stratum plexiforme externum) este compus din conexiunile sinaptice ce se realizeaza intre axonii celulelor vizuale (cu sferule terminale pentru celulele cu conuri si cu picioruse terminale pentru celulele cu conuri) si dendritele neuronilor bipolari.
Stratul nuclear intern (stratum nucleare internum) cuprinde corpul neuronilor bipolari,neuroni de asociatie (orizontali si amacrini) si corpul gliocitelor radiale (Müller). Nucleii neuronilor bipolari sunt situati in centrul stratului, aineuronilor orizontali in patura externa, iar ai glicocitelor radiale in patura interna.
Stratul plexiform intern (stratum plexiforme internum) este format din conexiunile sinaptice dintre axonii neuronilor bipolari si dendritele neuronilor multipolari.
Stratul ganglionar (stratum ganglionare) cuprinde corpul neuronilor multipolari.
Stratul fibrelor optice (stratum neurofibrarum) este format din axonii amielinici ai neuronilor multipolari, dispusi paralel cu suprafata retinei si din astrocite propoplasmatice. Axonii converg spre papila optica pentru a parasi globul ocular si a forma nervul optic.
Membrana limitanta interna (stratum limitans internum), formata din terminatiile prelungirilor interne ale gliocitelor radiale, apare ca o structura subtire si omogena.
Retina oarba. Prezinta o portiune ciliara si o portiune iriana, ce tapeteaza corpul ciliar si irisul. Are o structura foarte simpla, fiind formata din doua straturi celulare: unul intern si altul extern.
In portiunea ciliara, stratul extern continua epiteliul pigmentar al retinei vizuale, iar celulele stratului intern dau nastere unui sistem de fibre inextensibile si transparente groase de 2 - 5 mm, care se insera prin extremitatea lor pe regiunea ecuatoriala a cristalinului, formand
fibrele zonulei ciliare sau ligamentul suspensor al cristalinului. La nivelul proceselor ciliare, aceste celule participa la elaborarea umoarei apoase.
Stratul intern al retinei ciliare este acoperit pe fata sa interna de o membrana ce prelungeste in sens oral membrana limitanta interna a retinei.
Retina iriana cuprinde tot doua straturi ciliare, unul anterior care se metaplaziaza in elemente mioepiteliale ce formeaza muschiul dilatator al pupilei si altul posterior, epiteliul posterior al irisului, format din celule pigmentare.
Regiunile specifice ale retinei vizuale sunt reprezentate de pata galbena, ora serrata si papila optica.
- Pata galbena (macula lutea s. area centralis retinae) este o mica suprafata rotunda sau ovala, situata pe aria axului vizual al globului ocular, dorsal si lateral de papila optica. In centrul ei, la hominide, exista o mica depresiune, foveea centrala, la nivelul careia retina contine numai celule vizuale cu conuri, foarte inalte (70 - 85 mm) si foarte subtiri (2 - 2,5 mm). Spre marginea foveei, celulele cu conuri sunt mai putin lungi si mai groase si intercalate cu celule cu bastonas. Celulele stratului pigmentar sunt mai mici si subtiri. Stratul nuclear extern apare foarte gros, datorita abundentei celulelor vizuale cu con. Neuronii bipolari si multipolari sunt dispersati in cate un strat subtire. La nivelul petei galbene, fiecare neuron bipolar face sinapsa cu o singura celula vizuala cu con si cu un singur neuron multipolar, realizandu-se un plan functional continuu. Acest fapt nu permite o difuziune a excitatiilor, incat acuitatea vizuala este maxima, fata de celelalte regiuni ale retinei, unde o celula ganglionara primeste excitatii de la 100 - 2 000 celule vizuale. La anima lele domestice nu se intalneste depresiunea numita fovee centrala.
- Ora serrata este o zona de trecere intre retina vizuala si retina oarba. La nivelul ei celulele vizuale si neuronale sunt rare si atrofiate, iar celulele de sustinere sunt mai dezvoltate si tind sa ia forma unor celule epiteliale prismatice. Aceste celule prismatice intra in contact
prin fata lor externa cu celulele pigmentare care continua stratul pigmentar al retinei.
- Papila nervului optic sau pata oarba reprezinta o suprafata circulara prin care fibrele optice parasesc globul ocular, retina aparand atrofiata sau absenta. La acest nivel, sclerotica este ciuruita de orificii prin care trec fibrele nervului optic. In structura petei oarbe intra totalitatea fibrelor nervoase optice amielinice si cateva gliocite. In centrul papilei optice se gasesc artera si vena centrala a retinei.
Nervul optic cuprinde fibre optice formate din axonii neuronilor multipolari. In momentul in care trec prin orificiile sclerei, fibrele devin mielinice fara a prezenta o teaca Schwann bine structuralizata. Nervul optic este invelit de cele trei meninge (dura mater, pia mater si arahnoida), prezentand un spatiu arahnoidian care se continua cu spatiul subarahnoidian cerebral.
Intre celulele vizuale si diversele tipuri de neuroni din retina se realizeaza multiple si complexe conexiuni morfofunctionale. Astfel, s-au observat atat contacte directe intre axonii celulelor cu bastonase (terminate sub forma de sferule excavate) si axonii celulelor cu conuri
(terminali sub forma unor picioruse latite), cat si conexiuni prin intermediul neuronilor orizontali. In functie de conexiunile pe care le stabilesc, exista trei tipuri de neuroni bipolari: un tip ce stabileste contacte numai cu celulele vizuale cu conuri si doua tipuri ce stabilesc conexiuni cu un grup de celule vizuale cu bastonas. Neuronii bipolari ce intra in contact cu celulele vizuale cu conuri pot fi "neuroni trident", ale caror dendrite prezinta trei ramificatii, si neuroni bipolari cu "picior lat". In timp ce neuronii trident fac sinapsa cu o singura celula cu con, fiind prezenti mai ales la nivelul petei galbene, neuronii bipolari cu picior stabilesc sinapse cu 2 - 3 celule cu conuri. Neuronii amacrini stabilesc sinapse atat intre ei, cat si cu neuronii bipolari si multipolari. Neuronii multipolari sunt de doua tipuri, "trident" si "difuz".Neuronii multipolari de tip trident stabilesc sinapse cu un neuron bipolar trident, in timp ce neuronii multipolari difuzi stabilesc sinapse cu mai multi neuroni bipolari, aflati in conexiuni atat cu celulele cu bastonase, cat si cu celulele cu conuri.
Vasele retinei asigura nutritia retinei prin artera centrala a retinei si prin arterele ciliare. Artera centrala a retinei patrunde in globul ocular pe la nivelul papilei optice, dupa care se ramifica in 4 - 5 ramuri, ce ocolesc pata galbena, formand in grosimea retinei, doua plexuri in cele doua straturi plexiforme. Straturile externe ale retinei sunt lipsite de vase, nutritia realizandu-se prin difuziune din stratul corio-capilar. Artera centrala a retinei se ramifica diferit de la o specie la alta. La rumegatoare, suine, carnivore, rozatoare si primate, arterele retinale,
desprinse din artera centrala dau ramificatii arteriale si capilare in toata retina, realizand un model holoangiotic. Cabalinele si cobaiul prezinta un model paurangiotic, caracterizat prin vase mici, situate in jurul papilei optice. La leporide exista un model vascular merangiotic ce prezinta o artera centrala a retinei mica, care da ramificatii ce se limiteaza la o portiune redusa a retinei. La unele specii de pasari (liliac) si de mamifere (unele maimute), vasele retiniene nu sunt evidente, prezentand un model anangiotic. Ramurile ce provin din arterele ciliare se anastomozeaza, formand un inel scleral.
Venulele rezultate din retelele capilare conflueaza in vena centrala a retinei, care are un traiect invers arterei. Globul ocular este lipsit de vase limfatice.
1.4. Mediile transparente ale globului ocular
Sunt reprezentate de cornee (descrisa la tunica externa), umoarea apoasa, cristalin si corpul vitros.
Umoarea apoasa ocupa cele doua camere (anterioara si posterioara) ale globului ocular. Are aspectul unui lichid incolor, transparent, cu o compozitie chimica asemanatoare lichidului cerebrospinal. Nu coaguleaza si contine foarte mult acid hialuronic, in stare depolimerizata. Este produsa de procesele ciliare, dupa care trece o parte in corpul vitros,iar alta parte in camera posterioara, de unde prin orificiul pupilar ajunge in camera anterioara. Este resorbita la nivelul unghiului iridocorneean,de unde, prin venele ciliare anterioare, ajunge in formatiunile venoase ale sclerei. Indeplineste rolul de mediu refringent, participa la nutritia cristalinului si la reglarea presiunii intraoculare (de 23 mm Hg).
Cristalinul (lens) este o formatiune lenticulara, biconvexa, transparenta, elastica si incolora, situata aboral de iris, intre umoarea apoasa si corpul vitros. in structura cristalinului intra capsula, epiteliul anterior si fibrele cristaliniene.
Capsula sau cristaloida (capsula lentis) este o membrana transparenta, elastica, puternic retractila, cu structura lamelara. Lamelele contin fibrile de colagen, solidarizate de un material ce provine din lamina densa a membranei bazale. Cristaloida este mai groasa pe fata anterioara (8 - 12 mm), decat pe fata posterioara (2 mm).
Epiteliul anterior (epitelium lentis), dispus sub cristaloida anterioara este de tip simplu cubic; dispare la nivelul fetei posterioare. Celulele epiteliului prezinta un pol bazal, dispus pe cristaloida, si un pol apical, orientat spre fibrele cristalinului. La nivelul ecuatorului cristalinului,
celulele epiteliale devin alungite si se dispun in randuri concentrice. Regiunea ecuatoriala reprezinta zona de formare a noilor fibre cristaliniene prin modificarea celulelor epiteliale, incat la nivelul fetei posterioare nu exista epiteliu.
Fibrele cristaliniene formeaza tesutul propriu al cristalinuiui. Ele sunt celule ectodermice, alungite, care si-au pierdut prin adaptare functionala caracterul celular. Fibrele cristaliniene au forma de prisme hexagonale, sunt lipsite de nuclei si contin o citoplasma densa, fin granulara. Ele stabilesc jonctiuni prin interdigitatii, desmozomi si jonctiuni gap.Fibrele mature sunt lipsite de organite si ocupa centrul cristalinului,formand "nucleul". Fibrele mai tinere, situate periferic, formeaza corticala. Fibrele cristaliniene sunt incluse intr-o matrice amorfa cu rol de ciment intercelular. Aceasta matrice formeaza la polul anterior si posterior al cristalinului un fel de suturi (sutura lentis) sau aparate de sustinere, cu aspect de stele, pe care se insera fibrele. La om exista doua stele, una anterioara si alta posterioara, cu trei brate in forma de Y.In functie de insertie si directie, se descriu: fibre rectilinii, ce se intind de la un pol la altul; fibre principale sau mijlocii, curbe, inserate pe primele si dispersate in masa cristalinului; fibre superficiale, inserate pe unul din bratele stelei. La pasari, matricea cristaliniana, cu rol de ciment, se dispune punctiform pe ambele fete, formand o linie verticala pe fata anterioara, si orizontala, pe fata posterioara. La mamifere, dispunerea se realizeaza, ca si la om, in Y, cu orientare inversa pe cele doua fete.
Ligamentul suspensor al cristalinului sau zonula ciliara (zonula ciliara) mentine cristalinul in pozitie, fiind format dintr-un sistem de fibrile transparente, omogene, inextensibile. Fibrele (de 80 - 100 A diametru) se grupeaza, formand fibre mai groase sau mai subtiri, inserate pe de-o parte la nivelul corpului ciliar, iar pe de alta parte pe cristaloida, oral si aboral de ecuator. Fibrele ligamentului suspensor sunt elaborate de celulele nepigmentare ale retinei corpului ciliar. Ele mentin cristalinul in pozitie si ii transmit contractiile muschiului ciliar. In timpul contractiei, cristalinul se bombeaza, crescand puterea de convergenta.Cristalinul este lipsit de nervi si vase, incat nutritia cristalinului se realizeaza, prin difuziunea substantelor nutritive, din vasele de la nivelul proceselor ciliare si din umoarea apoasa. Cristalinul, impreuna cu corpul
ciliar, participa la acomodarea ochiului pentru vederea la distante variabile. Acomodarea la distanta joaca un rol mai mic la animalele domestice, unde muschiul ciliar este mai putin dezvoltat.
Corpul vitros, cu aspect de masa transparenta, cu consistenta gelatinoasa, ocupa camera interioara, delimitata intre cristalin si retina. Corpul vitros este un hidrogel (ce contine 98 - 99% apa), in care faza de dispersie este umoarea apoasa, iar faza dispersata este reprezentata de acid hialuronic si glicozaminoglicani acizi. Corpul vitros adera puternic la retina, la nivelul papilei optice si orei serrata. In regiunea orala prezinta o foseta patelara, determinata de fata posterioara a cristalinului, dela nivelul careia se intinde, in sens aboral, pana la nivelul papilei optice un rudiment al arterei hialoidiene, functionala la embrion, atrofiata la adult. La bovine, o reminiscenta a arterei hialoidiene poate aparea ca o cuta a membranei hialoide. La majoritatea animalelor domestice, corpul vitros este intim atasat capsulei posterioare cristaliniene, incat aceasta membrana trebuie pastrata intacta in timpul operatiei de cataracta, pentru a se evita prolapsul corpului vitros si colapsul camerei interne.
Ultrastructural, corpul vitros contine o retea fina de fibrile de colagen, care se condenseaza in regiunea periferica, formand membrana hialoida, ce ajunge in contact cu membrana limitanta interna. Corpul vitros joaca rolul unui mediu refringent, contribuie la mentinerea cristalinului in pozitie, contribuie la hranirea retinei ce impiedica dezlipirea acesteia.
Alte informatii referitoare la analizatorul vizual pe qgasesc.com| Contact |- ia legatura cu noi -| | |
| Adauga document |- pune-ti documente online -| | |
| Termeni & conditii de utilizare |- politica de cookies si de confidentialitate -| | |
| Copyright © |- 2024 - Toate drepturile rezervate -| |
|
|
|||
|
|||
|
|||
Proiecte pe aceeasi tema | |||
|
| |||
|
|||
|
|
|||