|






 |  |

| Biologie | Botanica | Chimie | Didactica | Fizica | Geografie |
| Gradinita | Literatura | Matematica |
Biologie
|
|
Qdidactic » didactica & scoala » biologie Sistemul fagocitar polimorfonuclear (PMN) |
Sistemul fagocitar polimorfonuclear (PMN)
Sistemul fagocitar polimorfonuclear (PMN)
PMNN sau granulocitele neutrofile sunt celule terminale, prezente la toate animalele cu aparat circulator, implicate in principal, in distrugerea MO patogene extracelulare. Datorita structurii lor si, in special, nucleului segmentat, sunt considerate ca fiind cele mai apte sa strabata spatiile inguste, cum ar fi cele dintre celulele endoteliale ale capilarelor, pentru a migra in tesuturile inflamate, prin diapedeza. La organismul uman si la majoritatea organismelor animale, neutrofilele sunt primele celule care intervin in raspunsul normal de aparare impotriva microorganismelor invadante. Neutrofilele sunt adaptate pentru a recunoaste, ingera si distruge majoritatea "invadatorilor straini" extracelulari, de dimensiuni mici.
Originea, diferentierea si caracterizarea generala a PMNN
Neutrofilele isi au originea in celula-stem pluripotenta din maduva oaselor. Celula stem, prin diviziuni repetate si diferentiere da nastere succesiv urmatoarelor stadii: mieloblast, promielocit, mielocit, metamielocit, PMN nesegmentate si segmentate.
Neutrofilul nesegmentat este prezent in maduva osoasa, dar in proportie mica si in sangele periferic. Neutrofilul segmentat, matur trece in sange, unde circula aprox. 10 ore, dupa care trece in tesuturi, unde mai traieste aproximativ 2-4 zile. In cursul procesului de maturare sunt intetizate si depozitate in granulatii diferite enzime cu rol degradativ si/sau antimicrobian. Granulopoieza sufera variatii de intensitate foarte mari si rapide. Numarul neutrofilelor creste masiv in cursul proceselor inflamatorii (leucocitoza), prin mobilizarea celulelor mature din maduva osoasa (90%).
PMN mature sunt repartizate in cele trei compartimente: rezerva medulara, neutrofilele
marginate si neutrofilele circulante. Neutrofilele constituie majoritatea leucocitelor sanguine circulante (aproximativ 70% la om). Numarul de neutrofile circulante creste, insa, rapid in cursul proceselor inflamatorii, prin mobilizarea celulelor aflate in maduva osoasa si intensificarea producerii lor.
Neutrofilele mature sunt celule cu nucleu segmentat si citoplasma bogata in granulatii, de unde denumirea de granulocite. Neutrofilele contin doua tipuri de granule, diferite prin dimensiune si continut:
a) granulatiile azurofile sau primare (A) au o forma sferica sau elipsoidala si diametrul de 500-800nm, contin enzime proteolitice active la pH neutru (elastaza, catepsina G, proteinaza 3), mieloperoxidaza (intreaga cantitate prezenta in celule), lizozim, hidrolaze acide (N-acetil b-glucozaminidaza, a-manozidaza). Efect: omorarea bacteriilor, degradarea diferitelor substante biologice;
b) granulatiile specifice sau secundare (B) sunt de dimensiuni mai reduse decat cele azurofile, de forma alungita, mai numeroase si se coloreaza palid (neutrofilie). Contin lactoferina (toata cantitatea), 90% din fosfataza alcalina celulara, restul de lizozim (muramidaza, dar are si un efect de perturbare a fenomenelor de oxidoreducere si implicit de blocare a proceselor de crestere si diviziune bacteriana), colagenaza etc. Sunt mobilizate brusc in perioada de formare a fagosomului cu care fuzioneaza si dispar din citoplasma. Continutul lor poate trece in spatiul extracelular (granulatiile azurofile fuzioneaza cu fagosomii mai tarziu si continutul lor ramane in fagosomi).
Rolul principal al PMNN, in stadiul de celule terminale, este de a fagocita si de a distruge bacteriile prin generarea de radicali liberi si eliberarea enzimelor stocate in granulatii.
PMN eozinofile. Sunt celule cu granulatii de tip special, care contin un spectru larg de enzime (fosfataze acide, ribonucleaze, catepsina, peroxidaza, sulfataze, arilsulfataze, b-glicuronidaze etc.). Prezente in numar mic in sangele circulant, prezinta miscari amiboidale, migreaza prin chemotaxie, fagociteaza si sufera degranulare. Activitatea lor microbicida este mai slaba decat a neutrofilelor. Fagociteaza eficient complexele antigen-anticorp, in special daca acestea contin IgE. Cresterea numarului peste normal (eozinofilia) poate fi determinata de productia crescuta in maduva sau de anumite categorii de antigene (de ex., parazitare) si mediatori.
Biomecanica neutrofilelor. Una dintre proprietatile cele mai evidente ale PMNN este mobilitatea, care le permite deplasarea activa in tesutul interstitial perivascular, pe endoteliul vaselor sanguine sau pe diferite suporturi in vitro (sticla, plastic). In mediu neutru, PMNN se deplaseaza la intamplare. Un rol important in mobilitate il are matricea proteica fibrilara, reprezentata de actina G care se transforma prin polimerizare in actina F (fibrilara) in prezenta ATP, miozina (cu activitate ATP-azica) si proteina de legare a actinei.
|
Etapele procesului de fagocitoza realizat de catre PMNN
1. Recunoasterea si opsonizarea. PMNN au receptori membranari, pentru molecule care declanseaza si intensifica fagocitoza, respectiv pentru opsonine: receptori pentru fragmentul Fc al Ig-lor si receptori pentru componentul C3b al sist.complement (SC). Opsonizarea (gr. opsonein = a pregati alimentele) este procesul de "sensibilizare" specifica sau nespecifica a bacteriilor, a altor celule sau antigene particulate fata de actiunea fagocitelor. Mecanism : opsonizarea favorizeaza fagocitoza prin cresterea hidrofobicitatii suprafetei particulelor si reducerea fortelor de repulsie dintre bacterii si neutrofile, ambele incarcate electronegativ. Bacteriile foarte virulente, rezistente la fagocitoza prezinta structuri extraparietale cu caracter hidrofil (polizaharide capsulare, acizi teichoici, fimbrii, stratul proteic paracristalin etc.), din categoria adezinelor ce mediaza aderenta la substratul celular sensibil, dar se opun aderentei la membrana fagocitelor (efect antifagocitar). Bacteriile avirulente sau slab virulente au suprafete hidrofobe si sunt sensibile la fagocitoza.
Opsonizarea are un rol foarte important si in asa numita fagocitoza de suprafata, proces prin care MO infectante sunt ,,capturate,, intre mai multe neutrofile si suprafata tesuturilor sau, impreuna cu neutrofilele in cheagurile de fibrina. Procesul compenseaza lipsa Ac-lor din primele 5-6 zile de la infectie si se adauga la actiunea SC activat pe cale alternativa. Fagocitoza de suprafata este mai eficienta in tesuturi si mai putin eficienta in mediul lichid (lichid pleural, peritoneal, LCR etc.) si foarte important in cazul MO prea mari pentru a fi fagocitate de PMNN.
2. Chemotaxia. Celulele mobile ca neutrofilele sunt capabile de migrare orientata de-a lungul gradientilor chimici ai stimulilor. Migrarea extravasculara a neutrofilelor este cauzata de gradientii unor atractanti chimici care se realizeaza intre tesutul inflamat si vasele de sange. Migrarea directionata a neutrofilelor la om este mediata, in mare parte, de componente solubile ale sistemului complement (ca, de ex., C5a), care se formeaza ca urmare a clivarii proteinelor SC, ca o consecinta a activarii acestuia pe calea clasica sau alternativa. Alti chemoatractanti ai neutrofilelor sunt: produsi bacterieni (exemplu:N-formil-metionil peptide), produsi ai coagularii si fibrinolizei, lipide oxidate (exemplu: leucotriene B4) si produsi ai stimularii leucocitelor.
3. Aderenta neutrofilelor la endoteliul vascular si migrarea extravasculara. Neutrofilele din circulatia periferica formeaza doua subpopulatii de neutrofile. Una dintre acestea este compusa din neutrofilele care circula prin fluxul central in vasele de sange. Aceste celule circulante reprezinta aproximativ jumatate din neutrofilele sanguine la om. Celule marginate, reprezentate de restul de neutrofile sanguine se misca incet de-a lungul endoteliului vascular. In conditii normale, neutrofilele marginate nu adera la suprafata endoteliului vascular. Indiferent de cauza, unul dintre evenimentele timpurii care insotesc inflamatia acuta este o crestere a numarului de neutrofile marginalizate, aderente la endoteliul vascular. Aderenta este o conditie obligatorie pentru diapedeza neutrofilelor in compartimentul extravascular.
4. Endocitoza. Contactul celulelor bacteriene cu neutrofilele si opsonizarea determina activarea structurilor submembranare cu rol in biomecanica fagocitelor si formarea unor pseudopode, care se deplaseaza in jurul bacteriilor, altor particule sau celule fagocitabile, inconjurandu-le progresiv, pentru ca, in final, extremitatile distale ale pseudopodelor sa fuzioneze, ingloband bacteria sau particula intr-o vezicula fagocitara sau fagozom. Fenomenul de inglobare necesita deci, interactiunea secventiala intre liganzii opsonici, distribuiti pe suprafata bacteriei si receptorii fagocitari. Inglobarea este conditionata de o interactiune cu caracter continuu, si circumferential, care functioneaza prin mecanismul de « fermoar » (zipper-like mechanism).
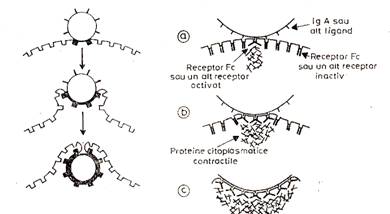
5. Degranularea si exocitoza. Dupa fagocitoza, granulatiile lizozomale fuzioneaza cu membrana vacuolei de fagocitoza si sunt descarcate enzimele lizozomale sau alti constituenti ai acestor granulatii (degranulare). Rezulta astfel structura numita fagolizozom. Granulele secundare fuzioneaza primele cu membrana fagozomilor.
Exocitoza. In anumite conditii, enzimele lizozomale sunt eliberate la exteriorul celulei, de ex., dupa moartea celulelor sau cand un neutrofil vine in contact cu diferite substante toxice. In alte cazuri, anumite substante patrund in interiorul sistemului vacuolar al celulei si produc ruptura membranei lizozomilor. Acest lucru duce la moartea celulei prin "perforare din interior" si la eliberarea enzimelor si a altor constituenti citoplasmatici (de ex., in cazul ingerarii de substante cristaline, ca uratul monosodic, siliciul).
Un alt mecanism prin care are loc eliberarea enzimelor lizozomale si a compusilor inalt reactivi ai oxigenului in spatiul ce inconjoara celulele intacte se pare ca se datoreaza eliminarii materialului lizozomal inspre spatiul extracitoplasmatic din fagozomii incomplet inchisi, dar in care granulatiile incep sa-si descarce activ continutul. Procesul are rol important in omorarea microorganismelor neinglobate, ci doar atasate de membrana fagocitului (fagocitoza de suprafata), cum este cazul infectiilor cu microfungi si paraziti (care interactioneaza cu eozinofilele). Actiunea enzimelor degradative este potentata de mieloperoxidaza si sistemele microbicide dependente de MPO, care determina omorarea MO parazite.
Sisteme bactericide active in PNM neutrofile
Procesul ingestiei MO opsonizate de catre neutrofile este necesar, dar, de obicei insuficient pentru omorarea acestor mocroorganisme. In timp ce mediul acid din interiorul vacuolelor fagocitare poate limita cresterea unor bacterii, distrugerea lor necesita produsi activi ai oxigenului molecular sau constituenti ai granulatiilor (enzimatici sau neenzimatici). Dupa ingestie, are loc o "explozie respiratorie" ce conduce la formarea de radicali anionici superoxid si a peroxidului de hidrogen, ca si la stimularea oxidarii glucozei pe calea HMP. Simultan, granulatiile citoplasmatice (lizosomii) fuzioneaza cu membrana fagozomilor. Produsii respiratiei celulare, impreuna cu constituentii granulatiilor actioneaza in interiorul vacuolelor fagocitare, omorand si degradand microorganismele ingerate.
In PMN neutrofile functioneaza doua sisteme bactericide: unele dependente, altele independente de O2.
I. Sisteme dependente de O2
sisteme dependente de MPO- enzima prezenta in granulatiile azurofile (rol esential, actiune rapida);
H2O2, format in metabolismul leucocitar;
halogenii (ionii de Cl-, I-) intra in vacuole prin membrana fagocitului sau odata cu particula
fagocitata; I- rezulta in ser prin aport alimentar, deiodarea hormonilor tiroidieni sau deiodarea neutrofilelor uzate, ca si din proteine bacteriene halogenate din peretele celular.
- MPO impreuna cu H2O2 si ionii de halogeni formeaza la pH acid un sistem microbicid foarte activ.
- sisteme independente de MPO (actiune mai lenta) - in cursul fagocitozei are loc reducerea univalenta a oxigenului, in prezenta NADPH si NADPH- oxidazei cu formarea:
- ionului superoxid - O2- (este prezent intrafagozomal, ca si extracelular; in citosol exista superoxid-dismutaza -SOD cu efect antioxidant, care protejeaza PMN);
- ionului hidroxil- HO-
- 1O2 - oxigenul singlet- o forma superioara energetic, care rezulta din starea de triplet, prin absorbtie de energie radianta sau chimica, cu durata de viata scurta si care reactioneaza cu substante organice cu dubla legatura, fiind letal pentru orice sistem biologic; revine la starea fundamentala prin disiparea energiei si emisie de lumina, caldura, reactii chimice; de altfel, detectarea fenomenului de emisie de cantitati mici de radiatii electromagnetice de catre PMN care fagociteaza sta la baza unui test de chemoluminiscenta, de estimare a capacitatii functionale a PMN (se utilizeaza tuburi sensibile fotomultiplicatoare).
II. Sisteme independente de O2
- lizozim (muramidaza) - mai activ pe bacteriile Gram pozitive;
- lactoferina- proteina chelatoare de fier, cu efect microbiostatic cand este saturata cu Fe; microorganismele se apara de actiunea LF prin producerea de siderofori, care competitioneaza cu LF pentru captarea Fe, necesar metabolismului microbian;
- proteine cationice bazice denumite generic fagocitine, care contin Lys cu rol in cresterea permeabilitatii peretelui celular bacterian de care se ataseaza; aminoacizii cationici ai acestor proteine cedeaza protoni pentru a forma punti cu componentele electronegative ale suprafetei bacteriene, astfel, proteinele cationice sunt fixate pe suprafata celulei bacteriene si afecteaza mecanismele de transport ale acesteia;
- hidrolaze acide, proteaze neutre;
histonele nucleare - eliberate in mediu in conditii fiziologice, dupa moartea celulelor, au efect bactericid.
Analiza comparativa a eficientei diferitelor sisteme a demonstrat rolul esential al sistemelor dependente de MPO, caruia nu-i rezista nici un microorganism, rezultand ca potentialul microbicid al neutrofilelor este in exces, redundanta fiind prezenta in amplitudinea unui sistem, ca si in diversitatea sistemelor; este benefica organismului, astfel ca daca un sistem este absent sau deficitar, celelalte il suplinesc.
Deficienta totala, globala a activitatii microbicide a PMN este rara si incurabila (0,025% din populatie este deficitara in MPO) si se manifesta prin infectii recurente, din primele luni de viata, la nivelul tegumentelor si mucoasei cailor respiratorii, sinusurilor - cea mai cunoscuta fiind granulomatoza cronica (PMN ingera bacterii, dar nu le poate omori, pentru ca nu pot converti O2 la intermediari microbicizi, lipsind NADPH-oxidaza care transfera electronii la O2).
* granulom = reactie inflamatorie ampla, datorata incapacitatii fagocitelor de a inactiva chemoatractantii si de a degrada Ag-le microbiene.
| Contact |- ia legatura cu noi -| | |
| Adauga document |- pune-ti documente online -| | |
| Termeni & conditii de utilizare |- politica de cookies si de confidentialitate -| | |
| Copyright © |- 2024 - Toate drepturile rezervate -| |
|
|
|||
|
|||
|
|||
Analize pe aceeasi tema | |||
|
| |||
|
|||
|
|
|||