|






 |  |

| Biologie | Botanica | Chimie | Didactica | Fizica | Geografie |
| Gradinita | Literatura | Matematica |
Biologie
|
|
Qdidactic » didactica & scoala » biologie Miscarea celulara |
Miscarea celulara
Miscarea celulara
Miscarea celulara reprezinta o caracteristica esetiala a materiei vii. Astfel, odata cu moartea celulei dispar si miscarile biologice, ramanand numai miscarea browniana, care este un fenomen fizic.
Miscarile celulare sunt extrem de variate, in general insa se pot distinge trei categorii, dupa cum urmeaza:
1. Miscarile din timpul contractiei musculare - se bazeaza pe interactiunea dintre actina si miozina.
2. Miscari care modifica raportul dintre celula si mediul extern:
miscarea ameboidala, se bazeaza pe interactiunea dintre actina si miozina;
miscarea flagelilor, se bazeaza pe interactiunea dintre microtubuli si dineina;
miscarea cililor, se bazeaza pe interactiunea dintre microtubule si dineina.
3. Miscari intracelulare, care nu modifica raportul dintre celula si mediul extern:
miscarile din diviziunea celulara, se bazeaza pe interactiunea dintre microtubule si dineina;
miscarile din microvili, se bazeaza pe interactiunea dintre actina si miozina;
curentii citoplasmatici, bazati atat pe interactiunea actina s imiozina cat si microtubule si dineina.
1. Miscarile care au la baza mecanismul de interactiune intre actina si miozina
Sistemul motil bazat pe interactiunea dintre actina si miozina are la baza urmatorul mecanism[1]: intre capetele globulare ale filamentelor de miozina si monomerii de actina din filamente se stabilesc punti transversale. Desfacerea progresiva a acestor punti si refacerea lor cu monomerul urmator de actina determina alunecarea filamentelor de actina si miozina unul peste altul si realizarea miscarii.
Contractia musculara
Este forma cea mai perfectionata de miscare care are la baza sistemul actina si miozina si apartine unor structuri specializate din muschii striati, netezi si muschiul cardiac.
In celula musculara striata se intalnesc organite specializate numite miofibrile, cu structura cilindrica si care sunt alcatuite din unitati contractile mai mici numite sarcomere.
In lungul miofibrilei se pot distinge benzi intunecate (benzi A, anizotrope) care alterneaza cu benzi luminoase (benzi I, izotrope).
Benzile intunecate, A, sunt traversate la mijloc de o banda subtire luminoasa numita membrana sau linia M, iar benzile luminoase, I sunt strabatute de o linie intunecata numita membrana sau linia Z. Un sarcomer este cuprins intre doua linii Z (Fig. 25). Benzile A contin in centrul lor numai filamente de miozina, iar la capete prezinta intercalate si filamente de actina.
Celula (fibra) musculara
Membrana Z Sarcomer Miofibrila
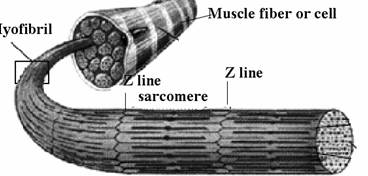
Fig. 25. Structura sarcomerului
Fiecare sarcomer contine la capete filamente de actina care se intrepatrund partial cu filamentele de miozina dispuse central.
Contractia musculara se produce prin glisarea filamentelor subtiri de actina pe langa cele groase de miozina, fara modificarea lungimii lor, glisarea avand ca rezultat scurtarea tuturor sarcomerelor si deci a intregii celule musculare. De fapt se scurteaza numai banda I, cea intunecata nemodificandu-si lungimea.
Procesul de contractie, de cuplare al actinei cu miozina se realizeaza numai in prezenta ionilor de calciu care sunt eliberati din reticulul endoplasmatic ca urmare a potentialului de actiune generat prin excitarea nervului motor al muschiului. Ionii de Ca2+ influenteaza proteinele asociate filamentelor de actina si anume troponina si tropomiozina, astfel incat sa faca posibila interactiunea actinei cu miozina. Energia necesara acestei interactiuni este furnizata de hidroliza ATP-ului. Relaxarea se face prin desfacerea simultana a tuturor puntilor dintre filamentele de actina si miozina (Fig. 26).
Filament de miozina Tropomiozina Filament de actina
![]()
![]()
![]()
![]()
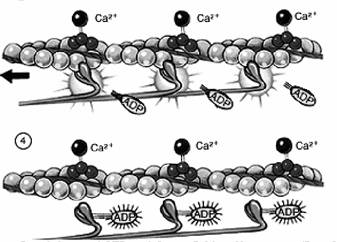
Fig. 26. Mecanismul molecular al contractiei musculare
Miscarile microvililor
Microvilii sunt expansiuni permanente de suprafata anumitor celule (enterocite, nefrocite, hepatocite, ovocite, celule senzoriale etc) implicate intr-o varietate de functii, incluzand absorbtia, secretia, adezivitatea celulara, mecanoreceptia etc..
Axa microvilului este alcatuita din aproximativ 40 de filamente de actina dispuse paralele si cu aceeasi polaritate. Filamentele de actina din microvili sunt legate din loc in loc cu ajutorul unor proteine adezive (fimbrina, vilina). Legaturile laterale ale filamentelor de actina cu plasmalema, in special in cazul microvililor enterocitelor, sunt realizate cu ajutorul unor " brate" formate din miozina si calmodulina (Fig. 27). La polul bazal, filamentele de actina se termina intr-o retea de filamente de miozina si spectrina, care formeaza butonul terminal, si care serveste la mentinerea pozitiei microvilului.
Spectrina Buton terminal Miozina Calmodulina si miozina Actina Fimbrina Vilina Filamente intermediare
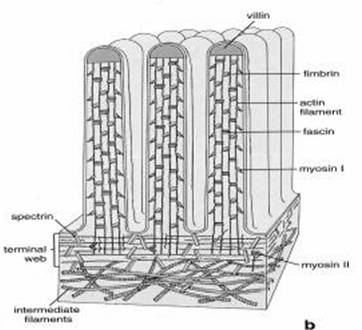
Fig. 27. Structura microvililor
Microvilii maresc suprafata de absorbtie a celulei, de exemplu la enterocite microvilii cresc de 25 de ori suprafata de absorbtie a substantelor nutritive. Datorita acestei functii vitale pentru enterocite, membrana microvililor este tapetata cu enzime care intervin in desfacerea substantelor nutritive complexe in compusi simpli, usor de asimilat (de exemplu: glicozidaza, care desface glucidele, este prezenta in concentratie foarte mare la suprafata microvililor enterocitelor). Prin urmare, microvilii nu doar maresc suprafata de absorbtie, ci maresc si numarul enzimelor cu rol digestiv prezente la suprafata celulara a enterocitelor.
La randul lor, microvilii sunt acoperiti de glicocalix, care participa fie la atasarea substantelor care urmeaza a fi absorbite, fie la protejarea acestora impotriva substantelor nocive. De asemeanea, glicocalixul microvililor poate constitui o alta locatie a enzimelor digestive.
Microvilii pot actiona ca si mecanoreceptori in nefrocitele situate in tubul contort proximal al rinichiului, unde sesizeaza debitul fluidului si converteste informatia, pe calea raspunsului biochimic, in reabsorbtie.
In cazul anumitor boli infectioase, in special a celor produse de Escherichia coli, datorita rearanjarii citoscheletului celulelor gazda, se poate produce distrugerea microvililor. Acest eveniment conduce la malabsorbtia substantelor nutritive, diaree osmotica persistenta, acompaniata frecvent de febra.
Uneori, distrugerea microvililor poate fi benefica, mai ales in cazul bolilor autoimune.
Lipsa congenitala a microvilor de la suprafata enterocitelor cauzeaza atrofia vilozitatilor intestinale, fiind, in general, o afectiune letala nou-nascutilor.
Miscarea ameboidala
Miscarea ameboidala este o forma de locomotie intalnita nu numai la protozoare, ci si la numeroase celule de metazoare, inclusiv la om.
Acest tip de locomotie prezinta o importanta deosebita deoarece este:
mecanismul prin care leucocitele parasesc vasele sanguine si se deplaseaza la locul infectiei in organism;
mecanismul prin care fibroblastele se deplaseaza la suprafata ranilor unde depun colagen si realizeaza cicatrizarea.
Miscarea ameboidala consta in emiterea si retractia continua a unor expansiuni membrano-citoplasmatice numite pseudopode. Formarea acestora se datoreaza unei treceri succesive ale citoplasmei din starea de gel in sol si invers. Astfel, formarea pseudopodelor incepe prin fluidificarea citoplasmei si scurgerea ei in pseudopode prin centrul acestora. La periferia pseudopodului, prin interactiunea dintre actina si miozina, are loc gelificarea, aderarea pseudopodului la substrat si trecerea corpului celulei in directia prelungirii.
In cazul leucocitelor, pseudopodele sunt mici si cu miscari lente, iar fibroblastele emit prelungiri latite, - lamelipode - care adera la substrat prin placi de adezivitate.
Curentii citoplasmatici
Curentii citoplasmatici sunt miscari intracelulare prin care are loc deplasarea organitelor si a componentelor in celula. Mecanismul molecular al curentilor citoplasmatici se pare ca se bazeaza in principal pe interactiunea actina si miozina. Anumite forme de curenti citoplasmatici, cum ar fi transportul organitelor celulare si a veziculelor sinaptice din neuroni, au la baza mecanismul microtubuli+dineina.
Miscari de tipul interactiunii actinei cu miozina se mai intalnesc si in formarea inelului tranzitoriu care apare la sfarsitul diviziunii celulare si duce la fragmentarea celulei initiale in cele doua celule fiice, precum si in cazul fibrelor de stres care apar la suprafata celulelor sub actiunea unor factori straini.
2. Miscarile care au la baza mecanismul de interactiune dintre microtubuli si dineina
Exista multe tipuri de celule care prezinta prelungiri cu caracter permanent mai mari decat microvilii si in a caror alcatuire intra microtubulii. Aceste organite sunt cilii si flagelii, care au un mecanism molecular al motilitati identic (Fig. 28).
La mamifere, cilii sunt prelungiri multiple la suprafata unor celule epiteliale din mucoasa nazala si traheala, unde dizloca si elimina particulele acumulate in secretia de mucus a acestor tesuturi, si din epiteliul mucoasei trompelor uterine, unde au rolul de a propulsa ovocitul din ovar in uter. Cilii care executa miscari ciclice in doi timpi: bataie (de inaintare) si revenire (inversa).
Flagelul este un organit de miscare caracteristic protozoarelor flagelate si gametilor masculi la vertebrate si care efectueaza miscari de insurubare, elicoidale si ondulatorii.
Structura cilului
Cilul este alcatuit din trei parti: portiunea libera, radacina si corpusculul bazal. Ultimele doua portiuni sunt plasate in citoplasma.
Portiunea libera este alcatuita dintr-o structura tubulara - axonema - care este invelita de o zona de citoplasma si de membrana celulara.
Axonema este formata din noua dublete de microtubuli periferici asezati in jurul a doi microtubuli centrali. Fiecare pereche este alcatuita din subfibra A, un microtubul complet cu 13 protofilamente si din subfibra B, care contine numai 10-11 protofilamente. Cei doi microtubuli centrali nu sunt contractili ci au rol in raspandirea impulsului de miscare, care porneste de la baza cilului. Alfa si beta-tubulinele sunt diferite in cele doua subfibre. Legatura dintre subfibrele unui dublet este intarita de o proteina asemanatoare cu cele din structura filamentelor intermediare, numita tectina.
De subfibra A se ataseaza o proteina numita dineina, care prezinta doua brate in forma de cleste, si care se gaseste in lungul microtubulilor la intervale egale, de 24 nanometri. Bratele dineinei ajung pana la subfibra B din dubletul vecin.
Dublete de microtubuli
In mentinerea structurii axonemei
participa trei tipuri de punti. Primul tip de punte este prezent
intre perechea de microtubuli centrali, care, in plus, sunt inconjurati de
de o structura fibroasa, numita teaca interna. Al doilea tip de punte este realizat de nexina, o proteina care
leaga dubletele periferice adiacente. Cel de al treilea ip de punte este
reprezentat de spitele radiare,
care pornesc radiar din perechea de microtubuli centrali spre fiecare
subfibra A din cele noua dublete periferice.
Nexina Spite radiare Teaca interna Tubuline Microtubuli Protofilamente Microtubuli periferici Subfibra A si B Corpusculul bazal Microtubulii centrali Plasmalema Bratele dineinei
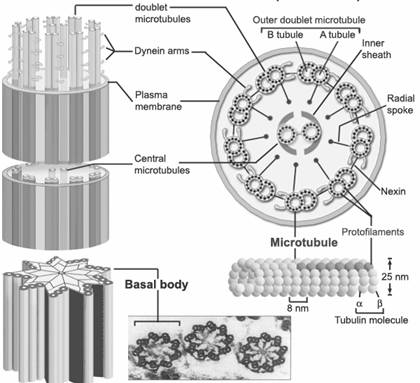
Fig. 28. Structura cililor si flagelilor
Structura flagelului
Structura flagelului este asemanatoare cu a cililor, dupa tipul 9 +2, dar in jurul axonemei se gaseste o coroana bogata de mitocondrii, care asigura energia (ATP-ul) necesara miscarii.
Mecanismul molecular de miscare al cililor si flagelilor este acelasi. Dineina este o proteina cu activitate ATP-azica. Energia rezultata din scindarea ATP-ului produce alunecarea dubletelor de microtubuli, unul in raport cu celalalt si incovoierea axonemei. Alunecarea dubletelor se realizeaza prin deplasarea puntilor laterale ce se stabilesc intre dineina si dubletul adiacent astfel incat are loc glisarea dubletelor unul fata de celalalt si indoirea cilului sau a flagelului.
Spitele si teaca interna a axonemei au rolul de a controla activitatea bratelor de dineina astfel incat sa se produca o unda coerenta de miscare a cilului sau flagelului. Absenta acestor formatiuni duce la imobilitate. De asemenea, absenta dineinei poate provoca sindromul cililor imobili care determina aparitia diverselor infectii ale cailor respiratorii sau sa cauzeze imobilitatea spermatozoizilor, conducand la sterilitate.
Centriolii si corpusculii bazali ai cililor si flagelilor
Centriolii si corpusculii bazali ai cililor si flagelilor sunt organite de coordonare a motilitatii si mai ales de asamblare a microtubulilor.
Studiile efectuate la microscopul electronic au relevat ca centriolii care formeaza centrosomul si corpusculii bazali au aceeasi structura, fiind ansambluri cilindrice alcatuite din 9 triplete de microtubuli periferici inveliti intr-o substanta densa si fara microtubuli centrali (9+0). Centriolii coordoneaza motilitatea celulara, in special in timpul diviziunii celulare, pe cand corpusculii bazali ancoreaza, asambleaza si coordoneaza miscarile cililor si flagelilor.
Ambele formatiuni au un rol important in polimerizarea tubulinei si formarea microtubulilor. Astfel, din rezervorul de tubuline al citoplasmei, centriolii induc formarea microtubulilor care formeaza fibrele fusului de diviziune iar corpusculii bazali duc la formarea microtubulilor axonemei. In acest fel se explica formarea si disparitia fusului de diviziune sau regenerarea unui flagel. Uneori, corpusculii bazali se trasnforma in centrioli, avand capacitatea de organizare a microtubulilor.
Centrosomul sau centrul celular este o structura mai complexa, alcatuita din doi centrioli asezati perpendicular unul pe altul si inconjurati de un material pericentriolar.
In perioada dintre doua diviziuni celulare (interfaza) centrosomul este plasat in apropierea nucleului si de la el pornesc microtubuli in toata citoplasma. In perioada S (de sinteza) a ciclului celular are loc dublarea centriolilor, iar in timpul mitozei cele doua perechi de centrioli se separa si migreaza la polii celulei, intre ei asamblandu-se microtubulii care formeaza fusul de diviziune.
Functia principala a centrosomului este aceea de centru organizator al microtubulilor. Materialul pericentriolar, alcatuit din proteine si ARN, detine un rol cheie in acest proces. Acest material determina polimerizerea tubulinei si asamblarea microtubulilor care au capatul - in centrosom iar capatul +, cu crestere rapida, este situat distal. Centrosomul are rol si in procesul de directionare a miscarilor ameboidale ale fibroblastelor si leucocitelor. In acest caz centrosomul se situeaza inaintea nucleului, pe directia miscarii, procesul de deplasare al celulei pe substrat necesitand un sistem de microtubuli. Un fibroblast la care se distrug microtubulii cu anumite substante chimice (colcemid) continua sa prezinte miscare ameboidala, insa nu directionata ci haotica. Un alt rol al centrosomului este si in definirea polaritatii celulei (in celulele epiteliale este plasat apical fata de nucleu).
| Contact |- ia legatura cu noi -| | |
| Adauga document |- pune-ti documente online -| | |
| Termeni & conditii de utilizare |- politica de cookies si de confidentialitate -| | |
| Copyright © |- 2026 - Toate drepturile rezervate -| |
|
|
|||
|
|||
|
|||
Lucrari pe aceeasi tema | |||
|
| |||
|
|||
|
|
|||