|






 |  |

| Biologie | Botanica | Chimie | Didactica | Fizica | Geografie |
| Gradinita | Literatura | Matematica |
Biologie
|
|
Qdidactic » didactica & scoala » biologie Organele hematopoietice si hemopoieza - maduva osoasa hematogena |
Organele hematopoietice si hemopoieza - maduva osoasa hematogena
Organele hemopoietice (organa hemopoietica) asigura formarea (generarea) si regenerarea continua a elementelor figurate ale sangelui, pe tot parcursul vietii organismului animal. Pe langa hemopoieza (haima = sange, poesis =a produce), aceste organe sunt implicate si in: - distrugerea elementelor sangvine (hemoclazie); - in metabolismul general si in functia de aparare imuna (prin producerea de anticorpi, fagocitoza, ultrafagocitoza etc.).
Organele hemo- si limfopoietice sunt reprezentate de: - maduva osoasa hematogena, ca organ mieloid, si de organele limfoide ( limfonoduli, splina, timus, complexe limfoepiteliale - amigdale, placi Payer, bursa lui Fabricius).
1. Structura generala a organelor hemopoIetice
Acestea au un plan de organizare comun, ce cuprinde: - scheletul de sustinere: - parenchimul; - reteaua vasculo-nervoasa.
Scheletul de sustinere este mai dezvoltatat in perioada embriofetala, dupa care regreseaza. Este reprezentat de capsula (capsula) si trabeculele (trabecula) ce se detaseaza de pe fata sa interna,realizand stroma organelor.
Capsula este formata din tesut conjunctiv dens, neorientat (dermocapsular). In maduva hematogena este reprezentata de endost, iar in nodurile (ganglionii) limfatice cuprinde si miocite netede. Capsula detaseaza trabecule care impart organul in loji, lobi si lobuli. Capsula si trabeculele formeaza un sistem de sustinere (stroma) pentru vase si nervi, mentinand forma organului. Spatiile delimitate de capsula si trabecule sunt ocupate de tesut conjunctiv reticulat sau citoreticul, care este format dintr-o retea de fibre de reticulina, dublata de celule cu aspect stelat, celule reticulare (reticulocytus). Pe masura definitivarii morfologiei, organele isi completeaza structura cu: histiocite (macrofage), plasmocite, mastocite, limfocite, granulocite, adipocite.
Scheletul de sustinere indeplineste in principal mai multe roluri: mecanic (prin stroma), citogenetic, metabolic si plastic. Imbatranirea organismului produce modificari si la nivelul stromei.
Parenchimul organelor hemo-si limfopoietice (textus hemopoieticus) este reprezentat de tesutul mieloid (textus myeloidus), in maduva rosie hematogena, sau de tesutul limfoid (textus lymphoideus) - in organele limfopoietice. Celulele parenchimatoase se dispun: - in cuiburi (exemplu in maduva hematogena), - pe zone (exemplu timus, ggl. limfatici), - in noduli (foliculi) etajati (exemplu amigdale, cecumuri, bursa lui Fabricius), sau difuz, in corionul mucoaselor.
La animalele tinere, celulele reticulare (reticulocitele) sunt cele mai numeroase. In activitatea maxima, tesutul mieloid sau limfoid este predominant, iar in faza de declin functional, predomina tesutul conjunctiv si adipos.
Reteaua vasculara are dublu rol, trofic si functional.Este formata exclusiv din capilare sangvine tipice sau atipice.
Inervatia cuprinde filete nervoase perivasculare.
2. MADUVA OSOASA HEMATOGENA
Maduva osoasa hematogena (medulla ossium rubra) este localizata in areolele oaselor spongioase si in canalul medular al oaselor lungi. Isi incepe activitatea hemopoietica in perioada embrio-fetala tarzie.Postnatal (posteclozional), in ea se formeaza eritrocitele, granulocitele si trombocitele.
Capsula este reprezentata de endost si detaseaza trabecule din fibre de colagen si rare fibre de reticulina.
Parenchimul sau tesutul mieloid (textus myeloidus) cuprinde grupe de celule polimorfe si policrome, situate in apropierea capilarelor sangvine. Celulele sunt elemente ale seriei eritrocitelor, trombocitelor si granulocitelor, aflate in diferite stadii de evolutie. Prezinta zone distincte in care se grupeaza celulele din aceeasi serie si noduli limfoizi care sunt resturi din perioada embriofetala si reprezinta sursa medulara a mononuclearelor.
Raportul dintre diferitele serii celulare, modificarile calitative si cantitative in structura populatiei sunt determinate de o serie de factori fiziologici, fizici, chimici, biologici, individuali (exemplu: maduva rosie este inlocuita de maduva galbena, apoi de cea cenusie).
Populatia celulara a tesutului mieloid provine din celula stem pluripotenta (numita in trecut hemocitoblast).
Reteaua vasculara cuprinde: - capilare tipice, provenite din ramificarea, modificarea arterelor de nutritie ale osului; - capilarele arteriolare se deschid in sinusurile sangvine (delimitate de celulele reticuloendoteliale, sprijinite pe o membrana bazala) ce formeaza teritoriul "colabat" si reprezinta portiunea initiala a formatiunilor venoase.
Circulatia medulara se caracterizeaza prin:- sistem capilar perfect inchis; circulatie discontinua (intermitenta) in teritoriul colabat; lipsa vaselor limfatice.
Inervatia este asigurata de fibrele nervoase mielinice (= receptori) si amielinice (= efectori), ce realizeaza retele pericapilare.
Maduva hematogena indeplineste mai multe functii: 1- de organ hematoformator total (in perioada embriofetala, cand produce elementele figurate din cele 5 serii) sau partial ( postnatal sau posteclozional, cand produce numai trombocitele, eritrocitele si granulocitele ); 2- de hemoclazie, distrugand eritrocitele si leucocitele imbatranite, cu ajutorul celulele reticulare nediferentiate si a celulelor macrofage; 3- functie metabolica, intervenind in metabolismul lipidelor, nucleoproteinelor, fierului, cuprului si cobaltului; 4-de fagocitoza, prin celulele cu rol fagic; 5- de secretie, producand proteine plasmatice; 6- de influentare ( interrelatie) cu tesutul osos ( de exemplu: proliferarea intensiva a tesutului mieloid produce cresterea porozitatii si slabirea rezistentei osului, iar procesele patologice din os provoaca modificari la nivelul maduvei.
La adult, maduva osoasa asigura in exclusivitate formarea diferitelor populatii de limfocite, cu toate ca nu este un organ limfoid.Poate restabili functiile sistemului imunitar in cazul transplantului efectuat la animale iradiate, dar netimectomizate. Celulele stem multipotentiale ce o colonizeaza provin din ficatul embrionar. In structurarea ei are un sistem vascular complex, in care sunt prezente toate tipurile de celule circulante: hematii, granulocite, mononucleare, megacariotice, limfocite, plasmocite si precursorii lor
3. HEMOPOIEZA
Hemopoieza (hemopoiesis) sau hemocitopoieza (hemocytopoiesis) este procesul de generare si inlocuire a elementelor sangvine.Este un proces continuu, care se desfasoara in tesuturile specializate (limfoid , mieloid) din parenchimul organelor hematoformatoare.
Se desfasoara in doua perioade ontogenetice: - hemogeneza prenatala si hemogeneza postnatala.
3.1.Hemopoieza prenatala
La mamifere, hemopoieza prenatala se desfasoara in trei stadii: - mezoblastic, hepato-timo-splenic si medulo-ganglionar.
In etapa mezodermica, elementele sangvine se formeaza extraembrionar, prin metaplazia celulelor mezenchimale ale insulelor WOLFF si PANDER din aria vasulara a sacului vitelin (fig. 1.).
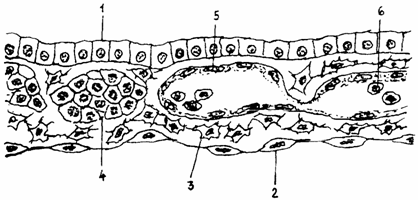
Fig. 1. Insule sangvino-vaso-formatoare:
1 - endoblast; 2 - splanchnopleura; 3 - mezenchim; 4 - Insula Wolff-Pander; 5 - endoteliu; 6 - Celule sangvine.
Insulele sunt aglomerari de celule, cu aspect sincitial, in care apar primele vase si primele celule sangvine ale viitorului organism (hemoangioblasti).
Celulele de la periferie isi pierd prelungirile,se aplatizeaza si se jonctioneaza, formand endoteliul capilar, ce delimiteaza vasul sangvin primitiv. Totodata ele secreta un lichid, care va deveni plasma sanguina.
Din peretele vascular se desprind primele celule mobile, ce plutesc in plasma sangvina, devenind elemente sangvine primitive sau hemoblasti. Unii dintre hemoblasti se incarca cu hemoglobina, devenind eritroblasti primitivi, care au un volum gigant, dar nu isi pierd nucleul si evolueaza pana la stadiul oxifil. Ei au o viata mai scurta decat eritrocitul adult. Acest lucru se realizeaza de timpuriu (dupa 14 - 15 zile ), datorita necesitatii crescute in oxigen,pentru realizarea proceselor de diferentiere celulara.
Etapa hepato-timo-splenica este caracteristica perioadei foetale.
Functia hemoformatoare revine parenchimului din trei organe: ficat, timus, splina, in care tesutul hemopoietic formeaza insule si infiltrate difuze.
In ficat - la foetusul de mamifere, hemopoieza are loc de timpuriu si dispare total dupa parturitie. In jurul sinusoidelor din mugurele hepatic, celulele reticulo-endoteliale se organizeaza in insule de proeritroblsti, eritroblasti si eritrocite. Apoi, apar megacariocitele si plasmocitele, iar mai tarziu granulocitele. La galinacee, ficatul posteclozional se mentine ca organ hematoformator (BOTAREL 1982 ).
Timusul prezinta o precocitate hemopoietica bazata pe capacitatea sa de a popula organele hematoformatoare periferice cu timocite.
Splina are initial doar functie eritroclazica,distrugand eritrocitele imbatranite ale altor organe, prin activitatea celulelor mezenchimale. Devine organ hemopoietic dupa hipertrofia si hiperplazia organului. In perioda fetala, splina produce majoritatea celulelor sangvine circulante. Paralel cu saderea si disparitia mielopoiezei, in splina se instaleaza limfopoieza, care persista ca functie definitiva.
|
Etapa medulara-ganglionara sau definitiva
Incepe prin intrarea in activitate a maduvei rosii hematogene si a ganglionilor limfatici in cea de-a 5-a parte a gestatiei, pe masura ce aceasta activitate regreseaza in ficat. Exista o perioada in care functia hemopoietica este exercitata in proportii aproximativ egale de maduva osoasa si de ficat-splina. In stadiul avansat al gestatiei, hemopoieza hepatica involueaza, iar hemopoieza medulara devine predominanta.
In perioada prenatala (preeclozionala ), precum si in primele zile dupa parturitie, apar organele cu functii hematoformatoare totale, ce produc si elibereaza elemente din toate cele cinci serii prezente in plasma: eritrocitara, granulocitara, limfocitara, monocitara si trombocitara.
La sfarsitul perioadei prenatale (preeclozionala) se manifesta o specializare a organelor hemoformatoare in organe mieloide si limfoide, insa fiecare din aceste organe poate reveni la capacitatea hemoformatoare totala si postnatal (posteclozional) ca o consecinta a "memoriei" pe care o pastreaza celulele mezenchimale "suse" (STEM).
In timpul ontogenezei, celule sangvine apar in urmatoarea ordine: 1) eritrocite; 2) granulocite; 3) trombocite; 4) limfocite si 5) monocite.
Diferente interspecifice
Intre diferitele specii de animale domestice exista diferente notabile cu privire la desfasurarea hematopoiezei prenatale (preeclozionale).
La canide, unde gestatia dureaza 60 - 65 zile, eritropoieza extraembrionara a fost obsevata la embrionul de 5 mm ( de 23 de zile) in membrana alantoida. Eritropoieza intraembrionara apare la embrionul de 10,5 mm, iar cea hepatica a devenit sesizabila dupa 40 de zile. In splina a fost observata dupa 45 zile, iar in maduva osoasa (la 48 de zile) incepe mielopoieza. In a 50-a zi de gestatie, maduva osasa prezinta tesutul hemopoietic bine organizat.
Formatiuni incipiente de timus au fost observate in a 25-a zi de gestatie, iar din a 40-a zi, timusul capata o structura apropiata de cea a cainelui adult. Nodurile limfatice (ganglionii) apar in ziua 48 de gestatie, incat la parturitie, placile Peyer si celelalte organe limfatice prezinta o structura aproape completa.
La feline, ficatul prezinta aglomerari de celule eritropoietice la foetusul de 35 zile, unde granulopoieza este putin reprezentata.
La bovine, embrionii de 4 mm prezentau eritrocite nucleate primitive. In ficat, primele insule eritrocitare au fost observate la embrionul de 4 - 8 mm. La acelasi stadiu de dezvoltare, s-au observat in ficat si primele megacariocite. La embrionul de 12 - 13 mm s-au observat primele neutrofile.
Hemopoieza hepatica atinge un maxim de dezvoltare la foetusii de 35 cm lungime (140 zile), dupa care descreste in intensitate, disparand la parturitie. Hemopoieza splenica incepe dupa 60 zile de dezvoltare intrauterina (foetus de 7 cm), atinge maximul la 200 zile (foetus de 60 cmm), dupa care descreste treptat pana la parturitie.
Hemopoieza medulara a fost observata in stadiul de 18 cm al foetusului, cand apar primele focare de eritopoieza, primele megacariocite, eozinofile si neutrofile. La 120 de zile de gestatiei (foetus de 30 cm), numarul neutrofilelor si metamielocitelor creste considerabil in maduva.
La pasari, dezvoltarea hemopoiezei difera de cea de la mamifere, intrucat pasarile au sacul vitelin dezvoltat. Rolul ficatul este lipsit de importanta pana la ecloziune. Inainte de ecloziune, maduva osoasa este implicata in producerea de eritrocite si granulocite, iar splina produce predominant granulocite, devenind predominant limfocitara numai dupa ecloziune.
3.2. Hemopoieza postnatalA
Hemopoieza postnatala sau posteclozionala se desfasoara in: organe mieloide ( maduva rosie hematogena ) si in organele limfoide primare (timus, bursa cloacala - Fabricius) sau secundare.
La canide, in timpul primei saptamani de viata postnatala, se mai observa insule hemopoietice in ficat, care dispar la 45 zile dupa parturitie.
Cu privire la formarea celulelor sangvine, exista patru teorii: unicista, dualista, trialista si pluralista.
Teoria unicista sau unifiletica, formulata de MAXIMOV si sustinuta mai recent de YOFFY, dupa care toate elementele sangvine provin dintr-o celula reticulara nediferentiata = celula STEM, celula susa multipotentiala sau HEMOHISTIOBLAST. Este aproape unanim admisa.
De asemenea, ACKERMAN sustine posibilitatea formarii elementelor sangvine din limfocitul mic.
Teoria dualista sau difiletica, emisa de NAEGELI, sustine formarea celulelor sangvine din doua tipuri de celule stem: unul pentru pentru granulocite si agranulocite, si altul, pentru eritrocite si trombocite.
Teoria trialista sau trifiletica, emisa de SCHILLING, sustine ca monocitele se formeaza din celule precursor, diferite de cele pentru granulocite si eritrocite.
Teroria pluralista sau polifiletica sustine existenta de celule stem distincte pentru fiecare tip de celula sangvina.
3.2.1. Celulele hemopoiezei
La baza formarii liniilor de celule sangvine sta o celula stem nediferentiata pluripotenta, diferita de celula reticulara. Dupa Berceanu -1977, celulele reticulare sunt celule stromale de sustinere, cu rol trofic, de stimulare a regenerarii si diferentierii, fara a fi celule hemopoietice.
Celula stem pluripotenta, numita in trecut hemocitoblast (hemocytoblastus), nu poate fi identificata morfologic, fiind asemanatoare unor limfocite mici, mature. Este o celula sferica, cu diametrul de 8 - 12 µm, cu o citoplasma fara granulatii, cu ribozomi liberi, cu putine vezicule si mitocondrii. Nucleul este sferic sau reniform, cu cromatina fin dispersata si nucleoli evidenti. Se gaseste in numar mare in maduva osoasa, dar este prezenta si in sagele circulant.
Celula stem pluripotenta genereaza nu numai celulele hemopoietice din maduva osoasa, dar si celule din alte tesuturi, fiind considerata o celula primitiva de origine mezenchimala. Ea este capabila de turnover sau de regenerare, incat poate mentine numarul optim de celule pe tot parcursul vietii organismului.
Prin mitoze succesive, celulele stem pluripotente asigura sursa de celule multipotentiale a maduvei osase hematogene.
O parte din aceste celule multipotentiale se angajeaza in diferentiere, generand celulele precursoare sau progenitoare angajate (commited) pentru fiecare linie celulara sangvina, care nu pot fi identificate pe criterii morfologice. Mentinerea celulelor precursoare in maduva este reglata prin mecanisme de tip feed-back de compartimentul stem pluripotent si de o serie de substante cu rol de hormoni, specifice pentru fiecare tip de celule sangvine (de exemplu: eritropoietina, granulopoietina etc.).
Dupa Junqueira-1998, celula stem pluripotenta prolifereaza si genereaza doua linii de celule orientate: - o linie din care se vor forma limfocitele, celule multipotentiale limfoide si o alta linie, celulele multipotentiale mieloide , din care se vor forma celulele mieloide, ce se vor vor dezvolta in maduva osoasa (granulocite, monocite, eritrocite si megacariocite). Fiicele celulelor multipotentiale isi reduc progresiv potentialiatea de diferentiere, transformandu-se in celule progenitoare, apoi in celule precursoare (sau blasti), care vor deveni celule mature.
Celulele progenitoare au un aspect general asemanator limfocitelor,fara a prezenta particularitati morfologice. Sunt mono- sau bipotentiale, se divid intens in maduva osasa si organele limfoide. Produc atat celule progenitoare, cat si precursoare.
Celule precursoare sunt monopotentiale, se divid intens in maduva si organele limfoide, generand numai celule mature. Incep sa prezinte particularitati morfologice de diferentire.
Celulele mature abunda in sange si in organele hemopoietice, nu se mai divid si prezinta caractere morfologice specifice. Astfel, celulele (stem) multipotentiale limfoide migreaza in organele limfoide, unde devin celule progenitoare, denumite celule formatoare de colonii limfocitare (lymphocite-colony forming cell = LCFC). Acestea genereaza limfoblasti (ca celule precursoare), care devin limfocite.
Celulele stem multipotentiale mieloide raman in maduva osoasa, unde devin celule progenitoare eritrocite, megacariocite, monocite si grnulocite (neutrofile, eozinofile, bazofile).
In hematopoieza postnatala, se parcurg patru etape: 1. diferentierea; 2. proliferarea; 3. cresterea si maturarea; 4. citodiabaza;
1. Diferentierea consta in transformarea celulelor progenitoare angajate (commited) in elemente cap de serie: proeritroblast - pentru seria rosie; mieloblast - pentru seria granulocitara; megacarioblast - pentru seria trombocitara; limfoblast - pentru seria limfocitara; monoblast - pentru seria monocitara.
2. Proliferarea (etapa de inmultire) - se realizeaza fie prin diviziunea homoblastica a celulelor progenitoare (cand din celula mama rezulta doua celule de acelasi tip), fie prin diviziunea heteroblastica - din care rezulta doua celule diferite.
3. Cresterea si maturarea incepe de la elementul "cap de serie", care este o celula precursoare, un blast care sufera importante modificari morfo-chimice si se maturizeaza in directia seriei respective.
4. Citodiabaza este etapa de eliberare a elementelor sangvine nou formate.
3.2.2. Eritropoieza
Pentru seria eritrocitara, celulele precursoare pot fi incluse in doua categorii: precursori neidentificati si precursori identificati morfologic. In categoria precursorilor neidentificati morfologic intra celulele formatoare de colonii eritrocitare (erytrocyte colony forming cell =ECFC), care sunt in fapt celule progenitoare. Precursorii identificati morfologic sunt reprezentati de: proeritroblaste (pronormoblaste), eritroblastele (normoblastele) bazofile, eritroblastele (normoblastele) policromatofile si reticulocite. Reticulocitele intra in circulatie si devin eritrocite adulte
Totalitatea globulelor rosii adulte si a precursorilor lor medulari constituie eritron-ul. Eritron-ul este in continua reinnoire, pierderea hematiilor din sangele periferic fiind inlocuita imediat prin activitatea maduvei hematogene.(fig.2)
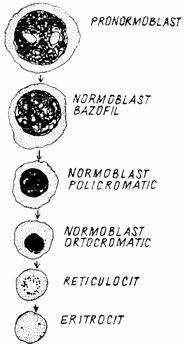
Fig. 2. Schema eritropoiezei
Proeritroblastul (proerythroblastus) este o celula, mare, sfeoidala sau ovalara, de 15 - 22 microni ( in functie de specie). Prezinta un nucleu voluminos, bogat in eucromatina, cu 1 - 2 nucleoli evidenti. Citoplasma, intens bazofila, prezinta o zona clara in jurul nucleului. Membrana periferica are numeroase invaginari, care dau nastere la vezicule de rofeocitoza, incarcate cu feritina. In proeritroblast se desfasoara intense sinteze de proteine, inclusiv de hemoglobina, care duc la cresterea volumului celular.
Se divide rapid prin mitoza, transformandu-se in eritroblast bazofil.
Eritroblastul bazofil (erithroblastus basophilicus) are un diametru de 15 - 20 microni. Nucleul ocupa 3/4 din celula, avand cromatina mai condensata, sub forma de grunji mari, in tabla de sah. Citoplasma, lipsita de granulatii, apare intens bazofila, incat eritroblastul bazofil este cea mai albastra celula medulara. Eritroblastele bazofile se divid mitotic, produc hemoglobina si se transforma in eritroblaste policromatofile.
Eritroblastul policromatofil (erythroblastus polychromatophilicus) are un diametru mai mic (9 - 16 microni) decat precedentul. Nucleul, picnotic, ocupa jumatate din celula, incat raportul nucleocitoplasmatic se egalizeaza. Nu se observa nucleoli. Citoplasma este caramizie, fara granule. Nucleul se poate fragmenta, generand corpusculii Jolly. Dupa fragmentarea si expulzia nucleului eritroblastului, celula se transforma in reticulocit. Unii autori descriu si un stadiu intermediar, de eritroblast oxifil sau acidofil (erithroblastus acidophilicus), datorita incarcarii citoplasmei cu hemoglobina (fig. 3.).
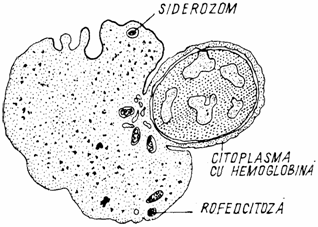
Fig. 3. Eliminarea nucleului din eritroblast.
Reticulocitul (erythrocytus reticulatus s. hemoreticulocytus) este o celula anucleata, un eritrocit tanar, cu diametrul de 3 - 10 microni (in functie de specie). In maduva, se matureaza in 24 - 48 ore. Inainte de a intra in circulatie, isi elimina organitele celulare, cu exceptia unor granule ribonucleoproteice, al caror continut in ARN confera bazofilie celulei. Prin coloratii postvitale cu albastru de cresil, aceste granule pot aparea ca granule izolate sau dispuse intr-o retea caracteristica reticulocitului.
Trecerea reticulocitelor in sangele circulant (citodiabaza) are loc prin diapedeza, celula emitand pseudopode, ce strabat peretele capilarului sinusoid medular. Din capilarele sinusoide medulare, reticulocitele trec in sangele circulant prin canale transendoteliale, unde sunt in proportie de 1 - 3% din eritrocite.
In apropierea capilarelor iau nastere insule eritroblastice, alcatuite din1 - 2 mcrofage, dispuse in centrul lor si inconjurate de de 1 - 2 randuri de eritroblaste (in numar de 2, 4, 8, 6 sau 32). Macrofagele emit prelungiri citoplasmatice ce se insinueaza printre eritroblaste. Toate eritroblastele dintr-o insula au acelasi grad de maturare, provenind dintr-o singura celula eritropoietica. Rolul asocierii dintre macrofage si eritroblaste nu este pe deplin clarificat. Se pare ca macrofagele intervin in fagocitarea si distrugerea nucleilor expulzati de eritroblaste, a eritroblastelor anormale, a eritrocitelor imbatranite sau patologice. De asemenea, macrofagele ar contribui la transferul feritinei catre eritroblaste, influentand activitatea eritroblastelor. Pe masura ce se formeaza si matureaza, globulele rosii ajung de la periferia capilarelor sinusoidale medulare (care formeaza teritoriul colabat, cu circulatie intermitenta) in centrul acestora, unde sunt antrenate de circulatia plasmei sangvine.
3.2.3. Granulocitopoieza (granulocytopoiesis)
Cuprinde procesele de formare a leucocitelor granulare. are loc in maduva osoasa hematogena dupa parturitie. Pentru formarea granulocitelor, se parcurge o etapa mitotica si o etapa postmitotica, rezultand trei compartimente medulare cu celule diferit maturate: 1-compartimentul celulelor stem pluripotente si unipotente; 2 - compartimentul de proliferare sau mitotic; 3- compartimentul de maturare si rezerva.
In compartimentul celulelor stem pluripotente si unipotente, se gasesc celulele stem multipotente care, sub actiunea unor stimuli adecvati, devin celule stem granulo-monocitare, cu morfologie neprecizata.
Compartimetul de proliferare sau mitotic cuprinde mieloblasti, promielociti si mielociti mari. Sunt prezente frecvente diviziuni mitotice. Mieloblastul se divide o singura data, promielocitul de 1 - 2 ori, iar mielocitul de 2 - 4 ori. Mielocitele sunt cele mai numeroase celule din acest compartiment, datorita numarului mai mare de generatii succesive.
In compartimentul de maturare si rezerva, celulele precursoare sufera o serie de modificari biochimice si morfologice, atat la nivelul citoplasmei, cat si la nivelul nucleului. Timpul de maturatie de la stadiul initial pana la celula adulta este de circa 5 zile. Dupa maturare, celule mai raman in maduva inca 2 - 3 zile. La unele specii, in rezervorul medular se gasesc mai multe granulocite decat in sangele periferic. La nevoie, are loc o mobilizare a granulocitelor din rezervorul medular.
De la celulele stem multipotente cu structura neprecizata, denumite celule formatoare de colonii granulocitare (granulocyte-colony forming cell = GCFC) se ajunge la granulocitul matur prin filiatia mieloblast, promielocit, mielocit, metamielocit si granulocit.
Mieloblastul (myeloblastus) provine din celula progenitoare angajata (commited) spre linia granulocitara. Este prima celula din linia granulocitara ce poate fi recunoscuta morfologic in maduva hematogena. Are o forma sferica sau ovoidala, cu un diametru de 16 - 20 µm. Nucleul este mare, ocupand 2/3 din celula, cu eucromatina predominanta si 1 - 3 nucleoli vizibili. Citoplasma este putina, bazofila (albastru deschis), dispusa uniform, cu cateva granulatii azurofile. Printr-o singura diviziune mitotica, fiecare mieloblast va da nastere la doua promielocite (fig. 4.).
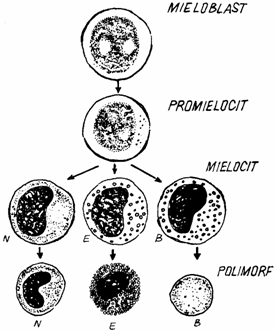
Fig. 4. Schema granulocitopoiezei.
Promielocitul (promielocytus) este usor mai mare decat mieloblastul. Nucleul apare sferic-ovoidal, bogat in eucromatina,dar si cu mici zone de heterocromatina. Nucleolii sunt slab vizibili. Citoplasma devine mai abundenta si mai intens bazofila, continand frecvente granulatii bazofile (granulatii primare), ce reprezinta lizozomii celulei. Traieste 24 - 48 ore, dupa care se divide mitotic, o singura data, generand doua mielocite.
Mielocitul (myelocytus) are un diametru mai redus (12 - 18 µm). Nucleul este ovalar la formele mari de mielociti sau usor incurbat la formele mici. Predomina heterocromatina, incat nucleolii nu mai sunt vizibili. Citoplasma apare ca un fond usor acidofil, bine circumscrisa in jurul nucleului, raportul nucleo-citoplasmatic fiind de 2/1. Pe langa granulatiile primare, apar si granulatii specifice secundare, dupa care se diferentiaza cele trei tipuri de mielocite: neutrofile (myelocytus neutrophilicus), eozinofile (myelocytus acidophilicus), bazofile (myelocytus basophilicus). Mielocitul are o viata de 24 - 48 ore, se divide de 2 - 3 sau 5 ori, generand celule mai diferentiate, metemielocitele, care trec in compartimentul nemitotic de maturare.
Metamielocitul (metamyeloccytus) are un diametru apropiat de cel al granulocitelor circulante. Nucleul prezinta o forma caracteristica, de rinichi sau potcoava, cu o indentatie adanca. Cromatina este densa si condensata in gramezi, separate prin benzi clare. Citoplasma este usor bazofila, cu organite slab dezvoltate. Activitatea de sinteza scade foarte mult, iar metamielocitele nu se mai divid. Numarul de granule primare se reduce, crescand, in schimb numarul granulelor specifice secundare (neutrofile, eozinofile, bazofile). Metamielocitii cu granule bazofile prezinta nucleu in forma de potcoava, putand avea cel mult o incizie mediana. Metamielocitul este stadiul cel mai tanar de granulocit diferentiat, putand sa apara in proportie redusa (de 1 - 5 % din granulocite) in sangele circulant.
Granulocitele nesegmentate (band cells) (neutrofile, eozinofile, bazofile) prezinta nucleul foarte curbat (reniform sau in potcoava), cu un inceput de formare a lobilor, fara separarea acestora. In sangele periferic, apare in proportie de 3 - 5% din granulocite. Numarul granulocitelor nesegmentate poate creste in sangele circulant in unele afectiuni, mai ales inflamatorii, cand se produce o stimulare a granulocitopoiezei.
Granulocitele adulte sau segmentate reprezinta ultimul stadiu de diferentiere, fiind granulocitele mature din sangele periferic. Eliberarea lor se face prin diabedeza, fiind controlata de un factor de eliberare (neutrophil releasing factor).
3.2.4. Monocitopoieza (monocytopoiesis)
Formarea monocitelor are loc in maduva hematogena, dintr-o celula progenitoare comuna pentru monocite si granulucite, celula granulomonocitopoietica (monocyte-granulocyte colony forming cell = MGCFC). Din aceasta celula ia nastere precursorul monocitar, monoblastul.
In maduva osoasa hematogena, monoblastul (monoblastus) nu poate fi deosebit de alte celule blastice, incat existenta sa ca celula intermediara intre celula granulomonocitopoietica si promonocit este pusa sub semnul intrebarii (fig. 5.).
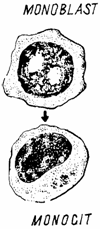
Fig. 5. Monocitopoieza - schema.
Promonocitul poate fi identificat dupa caracterele sale morfologice si functionale. Este prezent in numar redus in compartimentul de proliferare sau mitotic al maduvei osaose hematogene. Are un diametru de 10 - 20 µm si un raport nucleo-citoplasmatic mare. Nucleul este slab colorabil (palid), iar citoplasma apare bazofila, cu numerosi lizozomi, bogat reticul endoplasmic rugos si un redus reticul endoplasmic neted. Traieste 7 - 9 ore si se divide de 3 - 4 ori, dand nastere monocitelor.
O data formate, monocitele stationeaza foarte putin (circa 2 ore) in maduva osoasa, dupa care trec in circulatia sangvina, fara a se forma un rezervor medular, ca in cazul granulocitelor. In tesuturi, monocitele isi desavarsesc maturatia, devin active functional si apte pentru fagocitoza, schimbandu-si numele in macrofage tisulare. Numarul macrofagelor din tesuturi este incomparabil mai mare decat al monocitelor circulante, deoarece au o viata mai lunga (luni sau ani) si se pot multiplica local sub influenta unor stimuli adecvati. Distrugerea macrofagelor se face la nivelul lumenului intestinal si la cel al alveolelor pulmonare.
3.2.5. Limfocitopoieza (lymphocitopoiesis
In viata intrauterina, limfocitele s-au format in ficat, splina si maduva osasa (limfocitele B si T). Dupa parturitie sau ecloziune, ele se formeaza cu predilectie in organele limfoide primare, respectiv in maduva osoasa hematogena sau in bursa cloacala (limfocitele B) si in timus (limfocitele T). In masura mult mai mica, se formeaza si in organele limfoide secundare (limfonoduri, splina, placi Peyer). In maduva hematogena exista o celula stem limfoida multipotentiala, neidentificata morfologic. Dupa Junquiera -1998, aceasta migreaza in organele limfoide, unde devine celula progenitoare, cunoscuta sub numele de celula formatoare de colonii limfocitare (LCFC-lymphocyte-colony-forming cell). La randul ei, celula progenitoare incepe sa se diferentieze morfologic, se divide intens si se transforma in celula precursoare monopotentiala, cunoscuta sub numele de limfoblast (lymphoblastus) (fig. 6.).
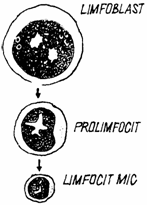
Fig. 6. Limfocitopoieza - schema.
Limfoblastul este prima celula a seriei, cu o morfologie ce ii permite sa fie identificata. Are un diametru de 16-20 µm, un nucleu sferic cu 1 - 2 nucleoli, raportul nucleo/citoplasmatic fiind in favoarea nucleului. Citoplasma este bazofila, cu multe organite si cateva granulatii azurofile. Ca aspect, se aseamana cu limfocitul activat. Se divide mitotic, dand nastere unei celule mult mai diferentiata, numita prolimfocit.
Prolimfocitul ajunge pana la un diametru de 15 µm. Nucleul contine o cromatina densa, cu grunji mari, si urme de nucleoli. Citoplasma, mai redusa cantitativ decat in limfoblast, este bazofila, cu organite rare si 5 - 6 granule azurofile,fine. Fara a se divide, prolimfocitul se transforma in limfocit circulant.
Prin metode de biochimie, enzimologie si marcare cu izotopi radioactivi, s-a demonstrat ca limfocitele cu acelasi aspect morfologic pot fi diferite functional. Astfel, o celula limfocitara cu aspect blastic poate fi un limfoblast sau o celula matura transformata blastic. Limfocitele T (inainte si dupa stimulare), ca si limfocitele B (inainte de stimulare), sunt asemanatoare cu limfocitele mici. Acest aspect pledeaza pentru utilizarea unei terminologii bazata pe criterii functionale, respectiv de imunoblasti si imunocite (virgine, sensibilizate,efectoare, cu memorie). Plasmocitul se diferentiaza diferentiaza asemanator limfocitului, trecand prin stadiile de plasmoblast, proplasmocit si plasmocit. Astfel, plasmoblastul are talia de 20 - 25 µm, un nucleu sferic, situat excentric, cu cromatina reticulara, grunjoasa si 1 - 2 nucleoli. Citoplasma este abundenta, bazofila, cu nuante violocee. Proplasmocitul seamana cu plasmoblastul, dar este de talie mai mica si nu are nucleoli. Plasmocitul scade in dimensiuni pana la 10 - 15 µm. Nucleul este sferic, situat excentric, cu structura grosolana, cu blocuri de cromatina dispuse radiar, in spita de roata. Citoplasma devine intens bazofila, albastra ca marea, usor violacee, cu o zona perinucleara mai clara (halou). Contin frecvente vacuole.
Pe frotiurile de sange pot aparea si unele aspecte morfologice atipice, legate de limfocite si plasmocite, cum sunt: -celula Turk, care are aspecte morfologice intermediare, intre limfocit si plasmocit; -celula Rieder, un limfocit cu nucleul foarte incizat (in trefla), intalnita frecvent in leucemia limfatica cronica; -umbra Gumprecht, rest de celula, cu nucleu amorf, liber, apare in leucemia limfatica cronica; - limfocitul monocitoid, cu talie mare, nucleu atipic, cu forme bizare, apare in leucemia limfatica acuta; - plasmocitele «flame», cu aspect flambat, citoplasma violet-roscata, apare in stari reactive si in mieloame; - celula Mott, un plasmocit ce contine imunoglobuline alterate, stocate sub forma de cristale sau vacuole.
3.2.6. Trombocitopoieza
Trombocitopoieza (trombocitopoiesis) are loc in maduva rosie hematogena, pornind de la celula stem multipotenta, din care se diferentiaza celula progenitoare (stem) unipotenta angajata (commited) spre formarea de trombocite, denumita celula formatoare de megacariocite (megakayocite- forming cell).
Aceasta celula da nastere unor celule precursoare diploide, care incep sa se divida mitotic intens, in momentul in care incepe procesul de diferentiere si maturare, generand megacarioblasti.
Megacarioblastul (megacarioblastus) este prima celula a seriei trombocitare care poate fi identificata in maduva rosie hematogena. Este o celula mult mai mare decat celelalte celule mieloide, ajungand la un diametru de 30 - 40 µm. Nucleul apare oval, uneori neregulat sau incizat, intens colorat. Citoplasma are o bazofilie pronuntata, cu zone mai intense, grunjoase (fig. 7.).
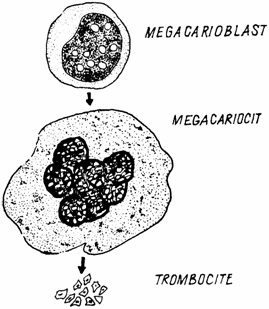
Fig. 7. Trombocitopoieza - schema.
Pe masura maturarii si aparitiei poliploidiei, megacarioblastul parcurge mai multe stadii de evolutie: I, II si III.
In stadiul I, megacarioblastul este o celula angajata, in care se divide numai nucleul, nu si citoplasma, proces denumit endoamitoza. Are 4 - 8 nuclei.
In stadiul II, apare megacariocitul bazofil sau promegacariocitul, care nu se mai divide. Acesta prezinta o poliploidie completa (4, 8, 16, rar 32 n). Nucleul este dens, tahicromatic, cu contur ovalar, cu incizuri si tendinta de impartire in lobi. Nucleolii nu sunt vizibili. Celula desfasoara o intensa activitate de sinteza, prezentand o citoplasma intens bazofila, cu numerosi ribozomi, poliribozomi, cu un bogat reticul endoplasmic rugos si complex Golgi.
In stadiul III, apare megacariocitul granular sau megacariocitul matur. Este o celula enorma, diametrul sau ajungand la peste 150 µm. Prezinta un nucleu segmentat, cu forme variate, cu lobi uniti prin filamente de cromatina, aparand un aspect de celula multinucleata. Unele celule sunt in mitoza, in timp ce altele sunt sunt in interfaza. Citoplasma apare impartita in zone bazofile si zone oxifile, cu granulatii pulverulente azurofile. Mai tarziu, toata citoplasma apare oxifila, cu granulatii fine, care pot fi dispuse in siruri paralele (aspect snuruit"), aspect ce anunta inceperea formarii trombocitelor. Pe masura ce se maturizeaza, talia celulei creste, iar forma devine ameboidala, cu pseudopode lungi.
Megacariocitul trombocitogen ia nastere prin maturarea megacariocitului granular. Are diametrul de 35 - 150 µm. Incepe sa descreasca in dimensiuni, prin picnotizarea nucleului, care este impins la periferie si prin condensarea citoplasmei. Citoplasma are un aspect pestrit, datorita aglomerarilor de granule azurofile. Organitele celulare apar mai reduse, in schimb sunt bine dezvoltate membranele, care vor forma membranele viitoarelor trombocite.
Membrana periferica emite numeroase pseudopode, care contin in axul citoplasmatic granule si numerosi microtubuli. Pseudopodele strabat peretele sinusoidelor medulare, trecand printre celulele endoteliale si ajungand in sangele sinusoidelor medulare. Din aceste pseudopode se desprind trombocite premature, care vor deveni trombocite mature, de forma discoidala, cu benzi microtubulare marginale.
| Contact |- ia legatura cu noi -| | |
| Adauga document |- pune-ti documente online -| | |
| Termeni & conditii de utilizare |- politica de cookies si de confidentialitate -| | |
| Copyright © |- 2026 - Toate drepturile rezervate -| |
|
|
|||
|
|||
|
|||
Lucrari pe aceeasi tema | |||
|
| |||
|
|||
|
|
|||