|






 |  |

| Biologie | Botanica | Chimie | Didactica | Fizica | Geografie |
| Gradinita | Literatura | Matematica |
Biologie
|
|
Qdidactic » didactica & scoala » biologie Organitele celulare |
Organitele celulare
ORGAniTELE CELULARe
Organitele celulare sunt diferentieri membrano-citoplasmatice care se formeaza pe o anumita treapta de evolutie a celulei, odata cu cresterea complexitatii morfostructurale.
In celula exista organite generale, comune tuturor tipurilor de celule ca de exemplu: reticulul endoplasmatic, mitocondriile, complexul Golgi, lizozomii, ribozomii, microtubulii, peroxizomii, centrul celular si organite specifice, intalnite numai la anumite celule cu functii speciale, de exemplu cilii si flagelii.
Unele organite celulare sunt delimitate de citomembrane (mitocondriile, reticulul endoplasmatic, lizozomii, complexul Golgi) iar altele sunt granulare (ribozomii) sau filamentoase (microtubulii si centrul celular), fara a fi invelite de citomembrane.
RETICULUL ENDOPLASMATIC
In toate celulele eucariote, cu exceptia eritrocitelor, s-a semnalat existenta unui complex de membrane intracelulare, organizate in tuburi, vezicule si cisterne, care strabat citoplasma intre nucleu si citoscheletul membranei celulare. Deoarece intinderea acestor formatiuni membranare se limiteaza numai la portiunea mai fluida a citoplasmei, acest organit a fost numit reticul endoplasmatic (lat. Reticulum = retea).
Primele observatii efectuate la microscopul optic au relevat existenta reticulului endoplasmatic, sub forma granulara sau lamelara, in celulele specializate in functia de sinteza proteica, cum sunt spre exemplu: hepatocitele, celulele acinoase ale pancreasului etc.
Studiile de microscopie electronica au evidentiat ultrastructura reticulului endoplasmatic[1], care este reprezentata de o retea extinsa de endomembrane dispuse sub forma de canale si sustinuta de citoschelet.
O caracteristica remarcabila a reticulului endoplasmatic este asocierea cu alte organite sau componente subcelulare. Astfel, anumite portiuni ale reticulului endoplasmatic prezinta ribozomi atasati (polizomi), alcatuind reticulul endoplasmatic rugos, iar alte portiuni, lipsite de ribozomi, formeaza reticulul endoplasmatic neted.
Reticulul endoplasmatic rugos este alcatuit mai ales din cisterne care continua membrana nucleara externa si are un rol esential in sinteza proteinelor. Prin urmare, acesta este mai dezvoltat in celulele cu sinteze proteice intense (celulele acinoase seroase, plasmocitele, unele celule endocrine cu rol in elaborarea hormonilor polipeptidici si glicoproteici).
Reticulul endoplasmatic neted se prezinta mai ales sub forma de tuburi sau canale interconectate care se intind spre periferia celulei pana la citoscheletul membranei si care constituie locul de sinteza, actiune si depozit al enzimelor cheie intervenind in procesele de sinteza ale lipidelor si steroizilor, in metabolismul glucidelor (contine enzima glucozo-6-fosfataza, care converteste glucoza-6-fosfat in glucoza, ca etapa in gluconeogeneza), in reglarea concentratiei de calciu, in reactiile de detoxifiere medicamentoasa etc. Din punct de vedere morfologic si functional, un aspect particular al reticulului endoplasmatic neted se remarca in celula musculara, unde, sub denumirea de reticul sarcoplasmatic, are rol in cuplarea excitatiei cu contractia.
Cele doua portiuni ale reticulului endoplasmatic nu sunt izolate una de alta, ele prezentand interconexiuni morfofunctionale (Fig. 29).
In precizarea compozitiei chimice si a functiilor reticulului endoplasmatic, o importanta deosebita au avut-o studiile biochimice efectuate dupa fractionarea celulei si centrifugarea diferentiata. Prin aceasta metoda, membranele reticulului endoplasmatic rugos s-au fragmentat si au fuzionat in vezicule mici numite microzomi, urmate de detasarea ribozomilor.
Studiile efectuate pe microzomi au precizat structura membranelor reticulului endoplasmatic, care sunt alcatuite din proteine (structurale, proteinenzime, hormoni) si lipide (predominand lectinele si cefalinele), in proportie aproximativ egala. Membranele reticulului endoplasmatic rugos contin in plus o categorie de glicoproteine transmembranare numite riboforine (ribofirina I si II), care au rol in legarea ribozomilor, in transferul prin membrana a lantului polipeptidic si in conferirea unei anumite forme cisternelor.

Reticulul endoplasmatic neted Reticulul endoplasmatic rugos
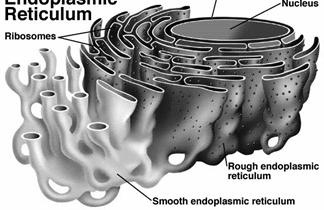
Fig. 29. Reticulul endoplasmatic
Proteinele din membrana reticulului endoplasmatic sunt glicoproteine care strabat complet stratul dublu lipidic, cu gruparile glucidice situate pe fata interna, lumenul reticulului endoplasmatic prezentand asemanari cu spatiul extracelular.
Lipidele din membrana reticulului endoplasmatic sunt in majoritate fosfolipide, cu acizi grasi foarte nesaturati, ceea ce confera o mare fluiditate membranei. Colesterolul se gaseste in cantitate redusa in membrane, insa este de doua ori mai mare in reticulul endoplasmatic neted.
Functiile reticulului endoplasmatic
1. Sinteza proteinelor de export si a proteinelor de membrana
Aceasta functie este realizata de reticulul endoplasmatic rugos, care apare bine dezvoltat in celulele cu functie de secretie. Lantul polipeptidic, in cazul proteinelor membranale poate ramane integrat in membrana reticulului endoplasmatic, poate sa treaca prin difuziune in membrana nucleului sau, pe calea veziculelor care se desprind din reticulul endoplasmatic, sa fie inclus in alte membrane.
2. Functia de biosinteza a lipidelor si a hormonilor steroizi, este realizata de reticulul endoplasmatic neted. Acesta este bine dezvoltat in celulele care sintetizeaza hormoni steroizi (corticosuprarenale, gonade) precum si in celulele absorbante din mucoasa intestinala, unde din acizii grasi, monogliceride si digliceride se sintetizeaza trigliceridele.
3. Functia de aparat circulator intracitoplasmatic este comuna atat reticulului endoplasmatic rugos cat si celui neted, care vehiculeaza in permanenta substante in toata citoplasma si in alte structuri, cum ar fi, spre exemplu: complexul Golgi si invelisul perinuclear.
4. Functia de transfer de substante intre citoplasma si lumenul reticulului endoplasmatic, realizata prin membrana reticulului atat prin difuziune cat si prin transport activ.
5. Functia de biosinteza a membranelor si organitelor celulare este una din cele mai importante functii ale reticulului endoplasmatic. In membranele reticulului endoplasmatic se sintetizeaza, proteinele si lipidele membranale si se elaboreaza enzimele destinate altor organite celulare cum sunt lizozomii si peroxizomii. Componentele sintetizate in membranele reticulului endoplasmatic pot ajunge la organitele "tinta" fie prin difuziune in planul membranei, fie prin vezicule de transport. Veziculele de transport se desprind de reticulul endoplasmatic prin inmugurire, incorporand o parte din lumenul si continutul reticulului endoplasmatic. Aceste vezicule se deplaseaza si fuzioneaza cu membranele organitelor tinta in care isi deverseaza continutul sau pot fuziona cu plasmalema eliberandu-si continutul la exterior. Reticulul endoplasmatic rugos este bine dezvoltat in neuroni care trebuie sa-si intretina o suprafata mare de membrane in prelungiri.
In celulele hepatice, reticulul endoplasmatic are rol important in degradarea glicogenului in glucoza si in procesele de detoxifiere. O serie de substante endogene sau exogene (medicamente, substante toxice) sunt metabolizate, neutralizandu-se efectele lor nocive si apoi eliminate prin intermediul reticulului endoplasmatic .
6.Rol mecanic. Prin sistemul de membrane si canale, reticulul endoplasmatic constituie un suport pentru citoplasma si pentru alte organite celulare. El realizeaza o compartimentare celulara si mentine forma celulei.
complexul golgi
Acest organit celular[2] se intalneste in toate tipurile de celule, cu exceptia eritrocitelor adulte (Fig. 30).
Pozitia in celula
Este diferita dupa tipul si activitatea citosolului. Astfel, in neuroni, complexul Golgi este dispus perinuclear, in celulele secretorii apare supranuclear situat intre nucleu si polul apical. In unele celule specializate (celulele foliculare din glanda tiroida) care prezinta dubla polaritate functionala, complexul Golgi se deplaseaza frecvent intre cei doi poli in raport cu activitatea metabolica predominanta a unuia sau a altuia. In majoritatea celulelor vegetale dar si in unele celule animale, complexul Golgi apare sub forma de dictiozomi (fomatiuni izolate sau grupate sub forma de tubuli anastomozati sau sub forma de cisterne).
Organizarea structurala si ultrastructurala
La microscopul luminescent, complexul Golgi se prezinta sub diferite aspecte: retea de canalicule anastomozate, vacuole de diferite marimi, bastonase.
Microscopul electronic a aratat existenta a trei elemente componente in organizarea complexului Golgi, respectiv:
microvezicule,
saci sau cisterne,
macrovezicule.
Toate aceste trei formatiuni sunt delimitate de citomembrane, de aproximativ 6-8 nm, cu aspect neted, fara ribozomi atasati pe suprafata lor, asemanatoare ca organizare, cu celalalte endomembrane lipoproteice, organizate in mozaic. In apropierea reticulului endoplasmatic, membranele golgiene se aseamana cu ale acestuia in timp ce cele mai indepartate, mai externe se aseamana cu plasmalema.
Sacii golgieni sunt formatiuni cavitare, usor curbate, plasate in apropierea nucleului. Numarul sacilor golgieni este in general de 5 - 11, ei fiind suprapusi si avand portiunea mijlocie mai ingusta si capetele mai dilatate. Structurile golgiene prezinta polaritate morfologica si functionala. Astfel, fata convexa a cisternelor este orientata spre nucleu si reticulul endoplasmatic rugos si se numeste fata "cis" (proximala sau imatura), iarfata opusa este concava si reprezinta fata "trans" (distala sau matura), orientata cel mai ades spre plasmalema.
Microveziculele sunt cavitati sferice delimitate de membrane, dispuse cel mai frecvent pe fata "cis" a saciilor golgieni. Microveziculele se desprind din membranele reticulului endoplasmatic invecinat, isi pierd ribozomii atasati si apoi fuzioneaza cu membrana cisternelor golgiene din apropiere.
Macroveziculele sunt formatiuni cavitare mari, care se desprind din partile periferice si de pe fata "trans" a sacilor golgieni. In majoritatea lor ele constituie vezicule de secretie care vor fi eliminate din celula prin exocitoza.
De la periferia cisternelor golgiene se formeaza si lizozomii primari. Structurile golgiene descrise isi au originea in membranele reticulului endoplasmatic sau, mai rar, din membrane preexistente ale sacilor golgieni.
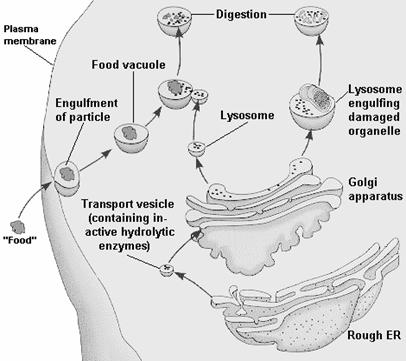
Reticul endoplasmatic Microvezicule Complexul Golgi -imagine la microscopul electronic- Nucleu Ribozomi Formarea de macrovezicule Vezicule secretorii Exocitoza Saci golgieni
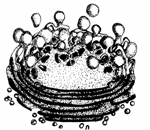
Fig. 30.Complexul Golgi si exocitoza (Mariana Sincai, 2000)
Functiile complexului Golgi
1. Functia de prelucrare si de transport a produsilor de secretie sintetizati in reticulul endoplasmatic. Complexul Golgi concentreaza produsii de secretie primiti de la reticulul endoplasmatic rugos, ii matureaza prin procese biochimice si ii inglobeaza in vezicule secretorii, care vor fi vehiculate spre plasmalema.
2. Biogeneza lizozomilor. Lizozomii reprezinta organitele prin care celula isi asigura moleculele fundamentale, atat pe baza reciclarii acestora din componente intracelulare, cat si prin preluari din afara celulei. Functia lor se bazeaza pe continutul bogat in hidrolaze acide, pe care celula si le produce si le directioneaza corect printr-o colaborare inalt specializata dintre reticulul endoplasmatic si complexul Golgi. Acest proces poarta denumirea de biogeneza lizosomala. El consta in biosinteza proteinelor lizosomale (enzime, proteine membranare lizosomale) care implica mai intai activitatea reticulului endoplasmatic, apoi pe cea a complexului Golgi pentru maturarea, sortarea si directionarea spre lizosomi a bagajului molecular specific. Prin urmare, lizozomii primari se formeaza ca urmare a interrelatiei structurale si functionale dintre complexul Golgi si reticulul endoplasmatic rugos, sistem numit GERL. Astfel, enzimele lizozomale sintetizate in reticulul endoplasmatic sunt aduse in cisternele golgiene pe fata "cis" de unde sunt trecute apoi, fie direct in veziculele de condensare, fie ajung pe fata "trans" unde se segrega de proteinele cu care au fost legate in sacii golgieni si genereaza vezicule lizozomale.
3. Functia de formare si circulatie a membranelor. Complexul Golgi primeste vezicule din reticulul endoplasmatic pe care le modifica (atat continutul cat si membranele) si apoi expediaza vezicule de secretie spre plasmalema unde, prin exocitoza, membranele veziculelor se integreaza in alasmalema. Suprafata celulei ramane insa constanta deoarece prin endocitoza se desprind din plasmalema vezicule care readuc in celula componente ale membranei pana la sacii golgieni.
|
La complexul Golgi pot ajunge proteine de membrana, receptori, si enzime care sunt modificate si refolosite, precum si molecule informationale din exterior (hormoni, catecolamine) care influenteaza metabolismul celular.
Fluxul endomembranelor poate fi alterat, atat din punctul de vedere al conexiunilor dintre formatiunile delimitate de endomembrane, cat si al manifestarilor functionale ale acestora. De asemeena, poate fi alterata si compozitia chimica a endomembranelor. Astfel de situatii pot fi datorate actiunii virusurilor sau transformarii maligne a celulelor.
4. Functia de generare a acrosomului. In spermatozoizi complexul Golgi genereaza supranuclear o vezicula care contine numeroase enzime litice, numita vezicula acrosomala. Enzimele pe care le contine lizeaza membranele celor doi gameti in timpul procesului de fecundare.
PEROXIZOMII
Peroxizomii sunt organite celulare specializate in producerea si descompunerea apei oxigenate[3].
Peroxizomii au forma sferica sau ovala, cu diametrul de 0,5 - 1 μm, fiind inconjurati de o membrana unica, de 6 nm grosime. Matricea este densa, omogena, continand, in unele celule si la unele specii, un cristaloid reprezentat de enzima uratoxidaza (Fig. 31). Acest cristaloid de uratoxidaza este prezent in mod normal in peroxizomii celulelor de sobolan si de alte specii, iar la om, in conditii normale, lipseste.
Membrana unica
Uratoxidaza Strat dublu lipidic
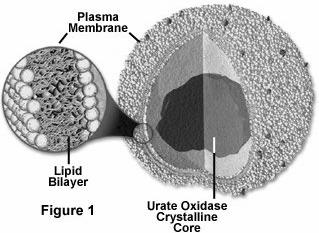
Fig. 31. Peroxizomii
Peroxizomii sunt foarte numerosi in hepatocite, nefrocite si macrofage. Aceste organite sunt generate prin inmugurire si fisiune din reticulul endoplasmatic neted sau din complexul Golgi. Unele enzime majore (catalaza, uratoxidaza) sunt sintetizate in citosol si apoi transportate in peroxizomii in curs de formare. Unii peroxizomi maturi raman atasati printr-o prelungire de reticulul endoplasmatic neted din care se formeaza.
Peroxizomii se pot autoreplica; enzimele si proteinele constitutive prezinta semnale tinta peroxizomale (peroxisomal targeting signals), care le atrage in peroxizomii nou formati.
Prin metode histochimice au fost evidentiati si microperoxizomi, de 0,2 μm (si care sunt lipsiti de cristalul de uratoxidaza), ei intalnindu-se in toate celulele mamiferelor.
Functiile peroxizomilor
In matricea peroxizomilor se gasesc enzime cu rol in producerea si descompunerea apei oxigenate.
Astfel, peroxizomii contin flavinenzime de exemplu: D-aminoacidoxidazele, L-α-hidroxiacidoxidazele care genereaza apa oxigenata prin utilizarea oxigenului molecular care se combina cu atomii de hidrogen din substrat dupa reactia:
RH2 + O2 →R + H2O2
Desi necesara in anumite reactii celulare si in distrugerea microorganismelor, apa oxigenata este tolerata de catre celule numai in concentratii mici. Excesul de apa oxigenata este convertit in apa prin actiunea catalazei.
La o concentratie mica a substraturilor cu hidrogen (RH2), catalaza va oxida apa oxigenata dupa reactia:
2H2O2 → 2H2O + O2
Daca exista un aport suficient de donori de hidrogen, reactia va avea loc astfel:
H2O2 + R'H2 → R + 2H2O.
Catalaza poate astfel oxida alcooli, fenoli, acidul formic, formaldehida etc., avand rol important in procesul de detoxifiere al celulelor.
Peroxizomii consuma in activitatea lor 10% din oxigenul ficatului. De asemenea, peroxizomii catalizeaza oxidarea acizilor grasi in acetil - CoA, care este transportata la mitocondrii pentru a intra in ciclul Krebs sau este utilizata in alte reactii de biosinteza.
Peroxizomii oxideaza acizii grasi cu lant lung de atomi de carbon pana cand lantul se scurteaza la 16 C, dupa care urmeaza oxidarea in mitocondrii, existand astfel o stransa cooperare intre peroxizomi si mitocondrii.
In ficat, peroxizomii prezinta urmatoarele functii: oxidarea excesului de acizi grasi, reduce excesul de H2O2, participa la sinteza colesterolului, participa in siteza acizilor biliari si reduce cantitatea purinelor aflate in exces (AMP, GMP) in acid uric.
Recent au fost descrise cateva implicatii ale peroxizomilor in anumite stari patologice. De exemplu, s-a constatat ca activitatea catalazei este foarte scazuta in ficatul animalelor cu tumori, iar in cazul tumorilor cu crestere rapida peroxizomii lipsesc. Numarul foarte redus sau absenta peroxizomilor a fost remarcata in sindromul cerebrohepatorenal Zellweger, intalnit la unii noi nascuti si care se manifesta prin hipotonie, acumulare de lipide in creier, hepatomegalie cu ciroza, chisti renali si anomalii scheletice, fenomene care duc la moartea individului in primul an de viata. De asemenea, o alta afectiune grava determinata de proasta functionare a peroxizomilor este si adrenoleucodistrofia (X-linked adrenoleukodystrophy - X-ALD), manifestata prin incapacitatea de a sitetiza acizi grasi, si care apare mai ales la baieti.
In unele maladii ca hepatita virala sau dupa administrarea indelungata de medicamente ce scad lipemia, peroxizomii cresc atat in dimensiuni cat si numeric.
CONDRIOSOMUL - MITOCONDRIA
Acest sistem in care fosforilarea si oxidarea sunt strans cuplate este cantonat intr-un organit celular special, numit condriosom sau mitocondrie. Cu exceptia eritrocitului matur, bazat in intregime pe glicoliza, in toate celulele se intalnesc un numar variat de mitocondrii, in functie de nevoile energetice ale celulei.
Mitocondriile sunt organite vacuolare, cu aspect granular sau filamentos[4] si care se intalnesc in toate celulele, cu exceptia eritrocitelor mature. Dimensiunile mitocondriilor sunt cuprinse intre 0,3 - 0,5 μm diametru si 10-20μm lungime. Mitocondriile se pot ramifica si fuziona, generand retele, sau se pot rupe in corpusculi mici, granulari, in conditii necunoscute inca. S-a observat ca lezarea celulelor duce la umflarea si fragmentarea mitocondriilor.
Numarul mitocondriilor variaza in functie de tipul de celula, intensitatea proceselor metabolice si gradul de dezvoltare al celulei, acest numar fiind in general estimat la 300 - 1000/celula.
Volumul total al mitocondriilor in majoritatea celulelor este de 5%, iar in celulele musculare de 30%.
La microscopul optic, in functie de activitatea celulei, condriosomul se poate prezenta sub trei forme:
- bastonase, o forma putin activa;
- filamente granulare, forma activa;
- granule dispersate, forma cea mai activa.
La microscopul electronic, indiferent de variatiile de forma, mitocondriile au aceeasi organizare de baza, ele fiind organite cavitare, ovalare sau sferice, marginite de o membrana dubla lipoproteica (Fig. 32).
Membrana externa a mitocondriilor este alcatuita din 60% proteine si 40% lipide, prezentand o permeabilitate mai mare comparativ cu membrana interna.
Membrana interna este plicaturata, prezentand o serie de falduri numite criste. Este alcatuita din 80% proteine si 20% lipide, fiind caracterizata printr-o permeabilitate mai redusa, dar cu un continut enzimatic mai bogat fata de membrana externa.
Intre cele doua membrane exista un compartiment mitocondrial extern.
Cristele pot avea o forma tubulara sau septala, pot fi simple sau ramificate, orientate perpendicular sau paralel cu axul longitudinal al organitului. Forma, numarul si dimensiunile cristelor variaza in raport cu tipul, activitatea metabolica si necesarul energetic al celulei. Astfel, cand activitatea metabolica a celulelor este intensa numarul cristelor creste.
Membrana interna delimiteaza un compartiment mitocondrial intern, in care se gaseste un continut amorf, cu o mare densitate, numit matrice mitocondriala.
criste
matrice granule ribozomi membrana interna membrana externa ADN mitocondrial
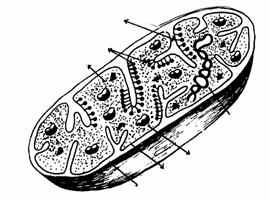
Fig. 32. Structura mitocondriei(Mariana Sincai, 2000)
De fata matriceala a membranei interne sunt atasate niste particule mici, prinse cu o tija fina. In matrice sunt inglobate granulele mitocondriale, ribozomii mitocondriali si genomul propriu, reprezentat de o molecula de ADN circular. Structura ADN-ului mitocondrial este asemanatoare cu a celui de tip bacterian. O mitocondrie contine aproximativ 2 - 6 molecule de ADN identice, care pot fi libere sau inlantuite. ADN-ul mitocondrial coordoneaza intreaga activitate a mitocondriilor, functionand independent de ADN-ul nuclear. In cazul in care genomul mitocondrial sufera mutatii, are loc alterarea structurala si functionala a mitocondriilor.
Modificarile morfofunctionale ale condriosomului sunt ciclice si se manifesta sub doua aspecte: starea de condensare si starea de gonflare (umflare).
In starea condensata, volumul compartimentului intern al mitocondriei se reduce, crescand densitatea matricei si numarul de criste. In starea gonflata volumul compartimentului intern creste, iar densitatea matricei si numarul cristelor scade.
Matricea mitocondriala contine pe langa proteine, lipide si glucide, o serie de vitamine (B, A, C) si ioni (K+, Ca2+ ,Mg2+) care sunt in cantitati variabile, dependente de metabolismul celular.
Condriosomul stabileste relatii cu nucleul, reticulul endoplasmatic, centrosomul, membrana celulara, etc.
O serie de factori patologici influenteaza morfofiziologia si numarul mitocondriilor. Astfel, inanitia si parathormonul in exces scad numarul de mitocondrii, la fel si acizii, alcoolii si hipertermia. Tiroidectomia determina cresterea numarului de mitocondrii.
Si pozitionarea mitocondriilor este corelata, in general, cu necesarul energetic al celulei gazda. Astfel, in celulele care sintetizeaza proteine, mitocondriile sunt situate in apropierea ribozomilor, in celulele angajate in transfer masiv de substante condriosomul este dispersat de-a lungul membranei celulare, iar in celulele musculare mitocondriile sunt concentrate in apropierea miofibrilelor.
Majoritatea cercetatorilor considera in prezent ca mitocondriile sunt la origine procariote ancestrale care au intrat in simbioza cu alte procariote gazda, generand celula eucariota. Aceasta ipoteza este sustinuta si de faptul ca mitocondria se divide independent de celula prin intermediul fisiunii binare, metoda a diviziunii celulei tipica procariotelor. Alte ipoteze privind originea mitocondriilor sunt: fragmentarea condriosomilor preexistenti sau inmuguriri ale membranelor reticulului endoplasmatic si ale plasmalemei.
Functiile mitocondriilor
Principala functie a mitocondriilor este aceea de a elibera energia continuta in moleculele organice simple (oze, acizi grasi, aminoacizi) si de a o inmagazina in molecule de ATP, pentru a putea fi utilizata prompt in numeroasele procese metabolice.
In mitocondrii au fost evidentiate peste 100 de enzime implicate in fosforilarea oxidativa. Oxidarea completa a unei molecule de glucoza la CO2 si H2O genereaza 36 molecule de ATP, din care doua sunt obtinute in timpul glicolizei anaerobe din citoplasma, iar 34 in mitocondrie. Influxul de substrat (piruvat sau acizi grasi) se realizeaza prin sisteme specifice de transportori. Efluxul de ATP produs de mitocondrii este cuplat cu influxul de ADP si se realizeaza printr-un sistem transportor specific. Astfel, se poate sintetiza ATP in mitocondrii numai daca exista ADP in citoplasma, respectiv daca celula foloseste ATP.
Tot in mitocondrii se desfasoara unele etape din sinteza hemului si a hormonilor steroizi.
De asemenea, ADN-ul mitocondrial este utilizat atat in depistarea unor boli genetice[5], pentru a identifica organisme antice care contin celule eucariote, cat si in medicina legala.
LIZOZOMII SAU CORPII LITICI
Lizozomii[6] sunt organite prezente in toate celulele, exceptand eritrocitele. Se gasesc in numar foarte mare in hepatocite si macrofage (aproximativ 200). Dimensiunea lizozomilor este cuprinsa intre 0,1 si 1,2 μm.
Studiile de microscopie electronica au relevat ca lizozomii (gr. lysis = disolutie, distrugere, topire, soma = corp) sunt organite vacuolare, delimitate de membrane lipoproteice si care contin numeroase enzime hidrolitice (peste 50 de tipuri de proteaze, nucleaze, lipaze, fosfataze, etc. - Fig. 33).
Aceste membrane lipoproteice permit enzimelor lizozomale sa actioneze, conform cerintelor, la un pH acid (4,5). Majoritatea lizozomilor sunt delimitati de o singura membrana, care protejeaza matricea citoplasmatica de actiunea enzimelor digestive. Insa, in nefrocite a fost semnalata prezenta unor lizozomi cu membrane duble, iar in macrofage s-au observat lizozomi a caror invelis prezinta spiculi radiari (lizozomi imbracati). De asemenea, membrana lizozomala, prin selectivitatea ei, controleaza substantele care patrund in lizozomi. Integralitatea membranelor lizozomale este influentata de o serie de factori: chimici, fizici si biologici.
Diferenta de pH existenta in lizozomi in comparatie cu cea din citosol se realizeaza cu ajutorul pompelor protonice.
O masura de protectie aditionala impotriva scurgerilor accidentale a hidrolazelor acide din lizozomi in citosol este reprezentata de sensiblitatea la pH a acestor enzime, care actioneaza mai putin eficient intr-un mediu usor alcalin (7,2).
In setul enzimatic sunt continute enzime cu rol important, dintre care:
- lipazele, care desfac lipidele;
- carbohidrazele, care desfac glucidele;
- proteazele, care desfac proteinele;
- nucleazele, care desfac acizii nucleici si acizii fosforici monoeterici.
Enzimele lizozomale sunt sintetizate in citosol si in reticulul endoplasmatic, care prin aditia manozei-6-fosfat, sunt atrase in lizozomi.
Membrana lizozomului Setul de enzime hidrolitice Proteine membranare glicozilate cu rol transportor Strat dublu lipoproteic
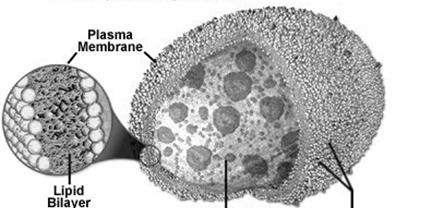
Fig. 33. Lizozomii
Actiunea digestiva a lizozomilor poate fi sistematizata in trei directii:
heterofagie - digerarea unor particule straine patrunse in celula;
autofagie - digerarea organitelor imbatranite si care se reinnoiesc in celula, a citomembranelor si a receptorilor;
crinofagie - digerarea produsilor de secretie in celulele endocrine, reglandu-se astfel nivelul secretiei acestora.
Actiunea de digestie celulara a lizozomilor, normala si patologica, se finalizeaza cu obtinerea unor produsi simpli (aminoacizi, monozaharide, acizi grasi) care trec prin membrana lizozomala in citoplasma, unde sunt reutilizati in procesele de biosinteza.
Pe parcursul activitatii lor, lizozomii trec prin mai multe etape marcate de o heterogenitate structurala (Fig. 34).
Lizozomii primari isi au originea in complexul Golgi si contin un set intact de enzime hidrolitice. Enzimele lizozomale, in cea mai mare parte glicoproteine, sunt sintetizate in reticulul endoplasmatic rugos, sunt aduse cu ajutorul veziculelor de transport in cisternele golgiene, unde sunt complexate si agregate in lizozomi primari.
In cazul heterofagiei, substantele straine celulei patrund prin endocitoza (fagocitoza sau pinocitoza). Prin endocitoza substantelor straine rezulta formatiuni veziculare delimitate de membrane, numite fie fagozomi (in cazul inglobarii de particule solide) fie pinozomi (in cazul inglobarii de macromolecule in solutie).
Lizozomii primari, prin chimiotactism, fuzioneaza cu heterofagozomii sau heteropinozomii, rezultand lizozomi secundari. Dupa fuziune se deverseaza in vacuola de digestie enzimele lizozomale specifice substratului, care vor efectua degradarea acestuia.
Reticul endoplasmatic rugos Fagozom Fagocitoza Vezicula transportoare
care contine enzime hidrolitice active Lizozom Fagozom Complex Golgi
Autofagie