|






 |  |

| Biologie | Botanica | Chimie | Didactica | Fizica | Geografie |
| Gradinita | Literatura | Matematica |
Biologie
|
|
Qdidactic » didactica & scoala » biologie Organele limfoide - timusul, bursa cloacala |
Organele limfoide - timusul, bursa cloacala
ORGANELE LIMFOIDE
Organele limfoide produc, stocheaza si "educa" limfocitele, care sunt principalele celulele efectoare ale apararii imune.
Totaliatea organelor limfoide alcatuiesc sistemul imun sau limfoid care cuprinde structurile si celulele din organismul animal care au capacitatea de a distinge selful de nonself si de a distruge sau inactiva substantele sau microorganismele straine ce patrund intr-un organism viu.
Componenta de baza a organelor limfoide este tesutul limfoid Tesutul limfoid este alcatuit din reticul si celule libere. Reticulul este format din fibre reticulare, celule reticulare si macrofage fixe. Celulele libere sunt, in marea lor majoritate, limfocite (aflate in stadii diferite de diferentiere si matuare), plasmocite si macrofage.
Exista 2 tipuri de tesut limofid: - tipul lax, in care predomina celulele reticulare; si - tipul dens, in care predomina limfocitele, care se organizeaza in noduli sau foliculi limfoizi.
Se deosebesc doua tipuri de organe limfoide: - primare (sau centrale) si secundare (sau periferice).
Organele limfoide primare apar primele in filogeneza si in embriogeneza. Sunt reprezentate de: timus si bursa cloacala ( Fabricius) si echivalentii ei la mamifere.
Organele limfoide secundare sunt reprezentate de: ganglioni (noduri) limfatice, splina, placi Peyer, apendice cecal, amigdale, vegetatii adenoide, noduli (foliculi) limfoizi, raspanditi difuzi in diferite mucoase. In filogeneza si in ontogeneza apar mai tarziu, dupa structurarea organelor primare. Sunt colonizate de celule diferentiate in organele centrale, celule ce ocupa teritorii specifice, denumite zone timus dependente", populate de limfocite "T" sau zone timus independente", ocupate de limfocitele B".
Au in structura lor o retea fina si complicata de vase sangvine si limfatice care creeaza posibilitatea ca limfocitele sa ia contact cu antigenele patrunse in organism sau in circulatie. Celulele sistemului imunitar au posibilitatea sa se recirculeze si sa se reintoarca in teritoriile specifice ( fenomen cunoscut sub denumirea de "HOMING"). In organele limfoide secundare, limfocitele au posibilitatea sa se diferentieze si sa se matureze in continuare, sub influenta stimulilor antigenici. Limfocitele pot sa se transforme in celule efectoare sau in celule cu memorie .
1. Organele limfoide primare sau centrale
Organele limfoide primare prezinta mai multe caractere generale:
1. apar mai de timpuriu in cursul dezvoltarii embrionare;
2. sunt centre de formare, diferentiere si diseminare in organism a limfocitelor specializate "T" sau "B";
3. in aceste organe, limfopoieza porneste de la celulele STEM multipotente si este independenta de stimularea antigenica. Limfopoieza este intensa inainte de nastere (cand animalul este mai putin expus la antigene), precum si la animalele "germ free";
4. extirparea organelor limfoide primare produce tulburari si deficiente ale raspunsului imun, cu atat mai grave, cu cat se efectueaza mai de timpuriu. Astfel, extirparea inainte de aparitia limfocitelor imunocompetente duce la suprimarea functiilor imunitatii specifice. Extirparea tardiva nu genereaza efecte grave, daca numarul limfocitelor imunocompetente nu este micsorat din alte cauze (iradiere, ser antilimfocitar,varsta inaintata).
5. dupa iradiere, organele limfoide primare pot fi repopulate numai cu celule STEM si nu cu limfocite diferentiate din limfonoduri (ganglioni), timus sau bursa;
6. dupa stimularea cu antigene, organele limfoide primare nu prezinta modificari citologice caracteristice raspunsului imun;
7. celulele produse de organele limfoide primare migreaza in organele limfoide secundare, unde isi desfasoara activitatea la nivelul unor compartimente functionale distincte.
1.1. Timusul
Timusul este un organ limfoepitelial, ce detine un rol esential in realizarea reactiilor de aparare imuna. La exterior, are o capsula de tesut conjunctiv, din care se desprind septuri , care impart parenchimul in mai multi lobuli.
Lobulul timic este unitatea morfofunctionala a timusului. Fiecare lobul are o zona corticala, periferica si o zona medulara, central. Zona corticala apare comuna pentru mai multi lobuli, trecand dintr-un lobul in altul (fig. 1.).
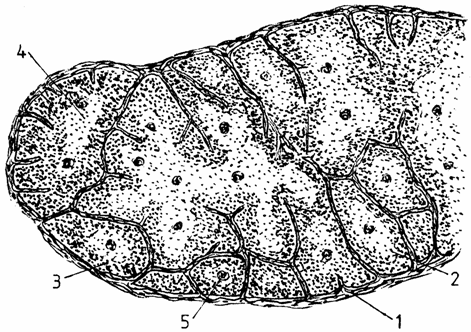
Fig. 1. Histostructura timusului
1 - capsula; 2- lobul; 3 - corticala; 4 - medulara; 5 - corpuscul timic.
Cortexul cuprinde o populatie densa de limfocite T, rare celule epiteliale reticulare, putine macrofage si limfocite mari. Celulele epiteliale reticulare sunt stelate, cu nuclei ovali, eucromatici. Ele jonctioneaza ele prin desmozomi si tonofilamente.
|
In raport cu structura si localizarea lor, se disting mai multe tipuri de celule epiteliale timice: celule doica, celule epiteliale dendritice din cortexul profund si celule epiteliale din medulara. Ele nu sunt celule limfoide, dar detin un rol cheie in proliferarea si diferenterea limfocitelor timice in curs de dezvoltare.
Celulele doica sunt celule specializate, localizate in stratul epitelial al cortexului extern. Ele se evidentiaza greu in timusul intact. Apar sub forma unor celule mari, care exprima pe suprafata lor antigene de clasa CMH II si cuprind limfocite corticale incluse in vezicule citoplasmatice. Unii autori considera ca ar fi timocite corticale (fig. 2.).
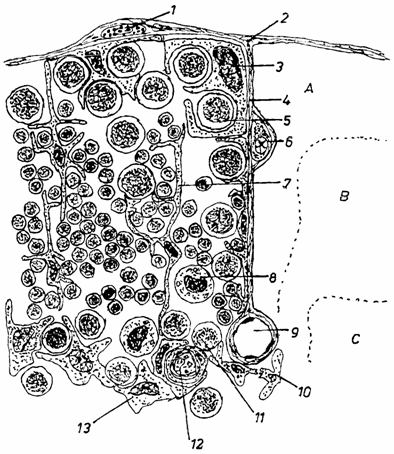
Fig. 2. Schema celulelor din componeneta timusului (dupa Hood-1984)
1 - mastocit extratimic; 2 - capsula; 3 - celula doica; 4 - sept; 5 - limfocit mare din cortex; 6 - plasmocit din sept; 7 - celula epiteliala dendritica corticala; 8 - macrofag; 9 - vas sangvin; 10 - celula dendritica interdigitata; 11 - limfocit mediu din medulara; 12 - corpuscul timic (hassal); 13 - celula epiteliala medulara.
Sunt considerate ca au origine epiteliala si rol in limfopoieza timica.
Celulele epiteliale dendritice se gasesc in cortexul profund. Prezinta numeroase prelungiri foarte ramificate (dendritice), care realizeaza o retea prin care trec limfocitele in drumul lor spre regiunea medulara. Ele delimiteaza spatii (lacuri) pline cu limfocite, care se gasesc la limita dintre corticala si medulara. Pe suprafata acestor celule si a prelungirilor lor se gasesc mari cantitati de antigene codificate ale CMH.
Celulele epiteliale din medulara au prelungiri lungi, care formeaza o retea laxa, in care se asigura maturarea limfocitelor T. Contin pe suprafata lor intreaga gama de antigene, codificata de genele CMH. Unele din aceste celule se grupeaza strans, se dispun concentric si formeaza corpusculii timici (HASSAL), relativ sferici, prin hialinizarea citoplasmei si dispunerea lor sub forma unor lamele concentrice in jurul unor zone centrale omogene eozinofile.
Corpusculilor HASSAL pot avea un rol secretor, probabil edocrin, si participa la distrugerea limfocitelor nematurate corespunzator, pentru a deveni imunocompetente.
In medulara lobulilor timici se gasesc doua tipuri de celule dendritice nefagocitare: - celulele reticulare denditritice si celule dendritice interdigitate, care contribuie la formarea unei tesaturi laxe. Ele provin din precursori medulari si sunt implicate in prezentarea antigenelor.
Macrofagele joaca rol de "sentinele", fiind dispuse pe ambele parti ale jonctiunii corticomedulare, in centrul curentului de limfocite.
Celulele limfoide din linia "T" sau timocitele formeaza populatia celulara dominanta. Ele ocupa reteaua formata de prelungirile ramificate ale celulelor epiteliale. Sunt mult mai numeroase in cortexul timic decat in medulara.
Rolul timusului. La inceputul vietii, timusul este un organ esential pentru maturarea imunitara normala a limfocitelor. La animalele adulte, timusul participa la amorsarea cu elemente a sistemului de aparare imuna de tip celular. Astfel, celulele epiteliale din timus furnizeaza seturile de antigene CMH si asigura "educatia" celulelor "T", pentru recunoasterea componentelor "SELF". Alte celule din timus, neidentificate inca, secreta mai multi hormoni peptidici implicati in maturarea limfocitelor T.
1.1.1. Organogeneza timusului
In filogeneza, timusul apare de timpuriu, inca de la pesti. La pasari si mamifere este singurul organ limfatic complet dezvoltat la nastere.
Timusul se dezvolta din epiteliul ventral al celei de a treia (la mamifere) si a patra (la pasari) pungi branhiale, in urma interactiunii dintre ectodermul branhial si endodermul faringian. Astfel, iau nastere doi diverticuli epiteliali ce se evagineaza in mezenchimul adiacent, se izoleaza de formatiunile faringiene, fuzioneaza in plan median , rezultand un conduct, care se orienteaza descendent, ajungand la locul de devoltare a organului ( fig. 3. ).
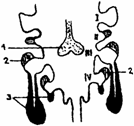
Fig. 3. Organogeneza timusului
1 - Tiroida; 2 - Paratiroidele; 3 - Timusul.
I - IV - Arcuri branhiale.
De timpuriu, timusul este populat cu celule stem pluripotente, care initial provin din sacul vitelin, apoi din ficat si din maduva osoasa. Aceste celule patrund in mezenchim si intre celulele epiteliului timic, alaturi de limfocite. Din celulele epiteliale se dezvolta citoreticulul timic. Corpusculii Hassal apar destul de timpuriu, fiind observati la foetusii suini de 60 zile.
1.2. Bursa cloacalA (fabricius)
Bursa cloacala sau diverticulul cloacal este un organ limfoepitelial specific pasarilor, cu aspect de punga, situat dorsal de proctodeum, cu care comunica printr-un canal bursal. Acest canal poate fi flancat de doua formatiuni bursale accesorii.
Peretele bursei este format din trei tunici: seroasa, musculoasa si mucoasa.
Seroasa este reprezentata de peritoneul local, care acopera diverticulul cloacal pe extremitatea sa cranio- dorsala.
Musculoasa este formata din fibre musculare netede, dispuse in doua straturi, longitudinal extern si circular intern.
Mucoasa ocupa cea mai mare parte din grosimea peretelui bursal, avand lamina propria ocupata, apropape in totalitate, de foliculi (noduli) bursali.
Mucoasa apare puternic cutata, formand pliuri groase, al caror centru este ocupat de foliculi (noduli), cu contur poliedric. Suprafata pliurilor este acoperita de un epiteliu simplu sau pseudoprismatic (Bacha-1990, Banks-1992) columnar. In acest epiteliu exista trei tipuri de celule columnare (I, II, III), in functie de granulele pe care le contin (fig. 4.).
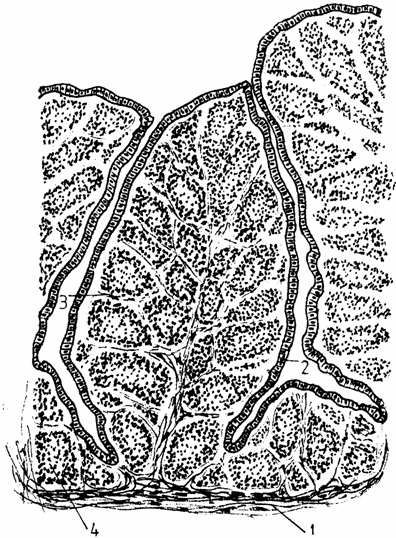
Fig. 4. Histostructura bursei cloacale
1 - peretele bursei; 2 - epiteliul simplu prismatic; 3 - corion;
4 - foliculi (noduli) bursali.
Aspectul de epiteliu simplu columnar este mai evident atunci cand acopera proeminentele. Acest epiteliu prolifereaza in profunzime, celulele epiteliale amestecandu-se cu celule din foliculi. Foliculii bursali pot avea doua aspecte: un aspect uniform, la foliculii primari, si cu doua zone, corticala si medulara, la foliculii secundari.
Foliculii (nodulii) bursali secundari apar dupa stimulari antigenice. Ei prezinta la periferia medularei un strat de celule epiteliale nediferentiate, cubice, si un strat de capilare. Aceste celule epiteliale cubice sunt implicate in activarea limfocitelor ce trec din corticala in medulara pentru a sintetiza anticorpi.
Corticala este separata de medulara printr-o membrana bazala. Zona corticala apare mai densa celular, cuprinzand numeroase limfocite, limfoblaste, celule in mitoza si numeroase macrofage. Vascularizatia zonei corticale este asigurata de arteriole, venule si capilare.
Zona medulara apare mai laxa celular si cuprinde celule nediferentiate, limfoblaste si limfocite. Vascularizatia zonei medulare este asigurata de o retea de capilare mari, dispusa imediat sub membrana bazala.
Rolul bursei consta in maturarea limfocitelor B, care vor secreta anticorpi, asigurand imunitatea de tip umoral. Initial, bursa are o structura pur epiteliala, fiind colonizata mai tarziu cu celule precursoare, venite din sacul vitelin sau din ficatul embrionar. Desi involueaza dupa ecloziune, isi mentine functiile prin care sunt maturate celulele B imature, aflate in circulatie. Eliminarea experimentala sau patologica a bursei determina aparitia unui deficit in functionarea limfocitelor B, cu atat mai sever, cu cat a trecut mai mult timp de la anihilarea bursei.
De asemenea, bursa produce un factor hormonal, bursopoietina, care, in vitro, induce diferentierea selectiva a limfocitelor B. Hood (1984) nu exclude posibilitatea ca si alte tesuturi hemopoietice sa participe, la pasari, la diferentirea limfocitelor B.
Limfocitele B parasesc bursa prin vasele sangvine care aprovizioneaza foliculii bursali, ajungand in tesuturile limfoide.
1.2.1. Organogeneza bursei cloacale (Fabricius)
Originea embrionara a bursei cloacale (Fabricius) suscita inca discutii. Primele cercetari considerau ca bursa are origine ectodermica, dezvoltandu-se din peretele superior al proctodeumului. Majoritatea cercetatorilor sustin originea endodermica a bursei, considerand-o ca o evaginare a endodermului pe fata dorsala a extremitatii caudale a intestinului posterior.
Primordiul bursal apare in zilele 4 - 8 de incubare, existand variatii legate de rasa si conditiile de incubare. Are forma unei proliferari, asezata pe fata dorsala a endodermului cloacal.
In epiteliul bursal apar grupe mici de celule mobile, cu origine limfoida, grupe care reprezinta fazele initiale de formare a foliculilor.
Mugurii epiteliali cresc rapid datorita mitozelor frecvente, astfel incat vor deplasa membrana bazala, infundand-o in corionul adiacent. Celulele epiteliale isi modifica aspectul initial si se diferentiaza in celule limfoide, sub inductia mezenchimului, cu care stabilesc raporturi morfofunctionale stranse.
Precursorii celulelor limfatice sunt celule stem pluripotente, ce provin din sacul vitelin, ficatul embrionar si maduva hematogena. Ele ajung pe cale sanguina in primordiul bursal, unde gasesc un mediu propice pentru proliferarea si maturarea lor. In apropierea ecloziunii, pliurile mucoasei bursale sunt bine dezvoltate, acoperite de un epiteliu, cu foliculi tineri in corion, lipsiti de zona corticala.
Dupa ecloziune, foliculii bursali prezinta o zona corticala, cu limfocite in diviziune rapida, impachetate dens, si o zona medulara, populata de limfocite dispersate si o retea de celule epiteliale.
Prin proliferarea rapida a limfocitelor apar cateva mii de foliculi limfoizi. In fiecare folicul limfoid sunt cuprinse circa 1 000 de limfocite B, care sunt diferite sub raportul capacitatii lor de a lega antigenele. Fiecare folicul contine limfocite B provenite dintr-o singura celula stem sau numai din cateva.
Bursa cloacala creste rapid in primele 3-4 saptamani dupa ecloziune, atinge dimensiuni maxime la 5 - 12 saptamani si involueza in perioada maturitatii sexuale, datorita influentelor hormonale.
| Contact |- ia legatura cu noi -| | |
| Adauga document |- pune-ti documente online -| | |
| Termeni & conditii de utilizare |- politica de cookies si de confidentialitate -| | |
| Copyright © |- 2024 - Toate drepturile rezervate -| |
|
|
|||
|
|||
|
|||
Referate pe aceeasi tema | |||
|
| |||
|
|||
|
|
|||