|






 |  |

| Biologie | Botanica | Chimie | Didactica | Fizica | Geografie |
| Gradinita | Literatura | Matematica |
Biologie
|
|
Qdidactic » didactica & scoala » biologie Transportul prin membranele celulare |
Transportul prin membranele celulare
TRANSPORTUL PRIN MEMBRANELE CELULARE
Membrana celulara este o bariera selectiva intre celule si mediul extracelular. Aceasta permeabilitate are drept consecinta faptul ca o serie de substante ca glucoza, aminoacizii si lipidele, care au intrat in celula, precum si o serie de metaboliti intermediari raman in celula, iar o alta gama larga de substante parasesc celula. Permeabilitatea selectiva a membranei celulare permite celulei sa-si mentina un mediu intern constant.
Transportul prin membranele celulare este grupat in doua mari categorii:
A. Transportul direct sau cu ajutorul unor proteine de membrana transportoare a unor ioni sau a unor molecule mici de substante organice {glucoza, aminoacizi).
B. Transportul prin includerea in vezicule de endocitoza generate de membrana celulara a unor macromolecule proteice sau a altor molecule organice mari.
1. TRANSPORTUL DIRECT SAU CU AJUTORUL UNOR PROTEINE
La randul sau, acest tip de transport transmembranar cuprinde doua categorii, dupa cum urmeaza:
Transportul pasiv, care se realizeaza fara consum de energie, de la un gradient chimic sau electrochimic ridicat spre gradientul scazut.
Transportul activ, care se realizeaza impotriva gradientului de concentratie, utilizand energia rezultata din hidroliza ATP-ului sau energia gradientelor chimice (sistemele antiport si simport).
Transportul pasiv
Transportul pasiv cuprinde mai multe tipuri de mecanisme dupa cum urmeaza (Fig. 4):
Difuziunea simpla
S-a constatat faptul ca patrunderea substantelor in celule este proportionala cu solubilitatea substantelor in lipide, masurata prin coeficientul de partitie al substantei intre ulei si apa. Cu cat acest coeficient este mai mare, cu atat substanta este mai liposolubila si patrunde mai repede in celule. Exista insa si exceptii de la aceasta regula, in care anumite molecule cum ar fi O2, CO2 sau molecule hidrofobe ca etanolul pot strabate stratul dublu lipidic fara ajutorul unor proteine transportoare.
Difuziunea facilitata
Plasmalema este impermeabila pentru multe molecule solubile in apa, precum: glucoza, nucleosidele, aminoacizii, H+, Na+, K+, Ca2+ etc. care sunt transferate prin membrane cu ajutorul unor proteine transportoare.
Difuziunea facilitata asigura pe de o parte o rata mai mare de transport a moleculelor prin membrana decat difuziunea simpla, iar pe de alta parte procesul prezinta o mai mare specificitate, fiecare proteina transportoare permitand transportul unui singur tip de molecule sau a unui grup foarte apropiat.
Referitor la difuziunea faciliatata de proteine transportoare amintim transportul ionilor prin intermediul ionoforilor (polipeptide produse de microorganisme).
Exista doua tipuri de ionofori: transportori mobili (carausi) si de tip canal.
Ionoforii din prima categorie sunt peptide ciclice care au la interior un spatiu polar in care sechestreaza un anumit tip de ioni. Un exemplu in acest sens este valinomicina care transporta K+ prin formarea in jurul ionului a unei sfere, cu partile hidrofobe ale aminoacizilor spre exterior si cu cei 6 sau 8 atomi de oxigen spre interior. Astfel se formeaza un complex valinomicina- K+ solubil in membrana dublu lipidica si care difuzeaza spre interiorul celulei.
Totusi, structura membranelor celulare cu un plan distinct hidrofob si altul hidrofil face foarte dificile aceste miscari de pendulare.
Ionoforii din a doua categorie preiau ionii de pe o suprafata a membranei, ii difuzeaza prin stratul dublu lipidic si ii elibereaza pe fata cealalta, prin formarea de pori. Un exemplu in acest sens este gramicidina, care este peptid liniar compus din 15 aminoacizi cu resturi laterale hidrofobe si care prin asocierea a doua molecule genereaza canale care strabat membranele celulare permitand cationilor monovalenti sa curga in sensul gradientului de concentratie. De retinut ca acesti dimeri sunt instabili, ei se formeaza si se desfac constant, timpul in care un astfel de canal este deschis fiind de o secunda, prin el trecand in acest timp 2x107 cationi. Alti ionofori de tip canal ca filipina, nistatina, amfotericina B formeaza pori doar in membranele cu continut de steroli (de exemplu membranele fungilor). De aceea sunt utilizate in practica medicala drept antifungice (antimicotice).
Ionoforii produsi de microorganisme sunt antibiotice, care pot constitui arme de aparare ale unor microorganisme impotriva altora, prin anihilarea potentialului de membrana.
De asemenea, ionoforii sunt folositi ca instrumente de investigare a multor procese in biologia celulara si moleculara. Printre altele, ionoforul A 23187 introduce Ca2+ si Mg2+ in celule, transportand in afara doi ioni de H+. Acest ionofor nu influenteaza potentialul membranar, doar creste concentratia intracelulara de Ca2+, fiind utilizat in studiul efectelor acestui tip de ioni.
O alta proteina canal bine cunoscuta este glucozopermeaza, cea care asigura patrunderea glucozei in celule conform gradientului de concentratie. Odata patrunsa in celula, glucoza este rapid fosforilata, in glucozo-6-fosfat, ceea ce face ca nivelul celular sa se mentina scazut si sa se permita continuarea importului acestui glucid in celula. Aceasta structura de proteina canal nu trebuie inteleasa ca un por permanent deschis care ar permite astfel si altor molecule asemanatoare glucozei sau mai mici (glicerolul) sa treaca nestingherit. In cadrul acestui tip de transport, glucozopermeaza este o proteina transmembranara care sufera modificari conformationale reversibile. Intr-o anumita stare conformationala ("pong"), locurile de legare pentru substanta ce trebuie transportata sunt expuse la exteriorul bistratului lipidic. In cealalta stare ("ping") aceleasi locuri de legare sunt expuse spre partea opusa a plasmalemei, iar substanta este eliberata. Acesta este mecanismul de "ping-pong".
De asemenea, in membrana celulelor au fost descoperite si alte proteine canal, numite aquaporine[1], cu rol in transportul transmembranar al apei.
C B A
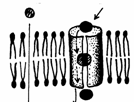
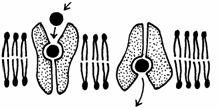
Fig. 4. Transportul pasiv al substantelor biologice: A - difuziune simpla directa,
B - difuziune prin proteine canal, C - difuziune facilitata prin proteine canal
(Mariana Sincai, 2000)
Un alt grup important de proteine canal ale membranei celulare sunt cele care asigura transportul ionilor.
Unele canale de transport sunt deschise in mod continuu, iar altele numai tranzitoriu. Acestea se numesc "canale de poarta" (Fig. 5). Unele canale se deschid la legarea unui ligand (mesager) pe un receptor si se numesc "canale cu poarta comandate de ligand". Alte canale dependente de potentialul membranei (voltaj) se numesc "canale cu poarta comandate de voltaj". Mai exista si canale proteice care se deschid ca raspuns la cresterea concentratiei intracelulare a unor ioni: de exemplu canalele pentru K+ se deschid atunci cand creste concentratia Ca2+ in citoplasma.
Exemple de canale cu poarta exista in functionarea jonctiunii neuromusculare, in care un impuls nervos produce contractia muschiului. Cel putin patru canale se deschid in mai putin de o secunda.
B Molecula transportata
Membrana
depolarizata Membrana
polarizata Gradient electrochimic
![]()
![]()
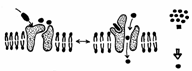
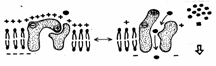
Fig. 5. A - Canale cu poarta comandate de ligand; B - canale cu poarta comandate de voltaj (Mariana Sincai, 2000)
Descresterea potentialului plasmalemei (depolarizarea) terminatiei neuronului deschide canalul pentru Ca2+. Cresterea concentratiei intracelulare de Ca2+ produce descarcarea veziculelor cu acetilcolina, care se leaga de receptorul acetilcolinei din plasmalema celulei musculare. Acesta este un canal cu poarta comandata de ligand; se produce deschiderea lui, ceea ce permite, timp de o milisecunda, difuziunea Na+ spre interior si a K+ la exteriorul celulei. Cum gradientul de concentratie al Na+ este mult mai mare ca cel al K+, influxul de Na+ depaseste efluxul de K+, deci se produce depolarizarea plasmalemei muschiului. Aceasta depolarizare deschide canalele pentru Na+, dependente de voltaj. Ceea ce produce o unda de depolarizare (potential de actiune), care se raspandeste pe intreaga membrana musculara. Ca rezultat al potentialului de actiune, se deschid canalele de Ca2+ in citoplasma. Aceasta crestere brusca a Ca2+ in citoplasma produce contractia miofibrilelor.
In cazul transportului pasiv prin membrana este necesar de mentionat ca aceste proteine canal asigura o permeabilitate selectiva, unii ioni putand sa treaca iar altii nu.
Gradientul de concentratie al ionilor si permeabilitatea selectiva a membranei creeaza o diferenta de potential intre interiorul si exteriorul celulei. Astfel, la interior celula este negativa electric, ca urmare a unei concentratii mai mari de anioni, iar la exterior este pozitiva, prin predominarea cationilor.
In ceea ce priveste cei trei cationi principali: Na+, K+ si Ca++, in interiorul celulei este mai mare concentratia ionilor de K+, iar in mediul extracelular a ionilor de Na+ si Ca++.
Gradientul ionic si potentialul electric al membranelor celulare este esential pentru multe fenomene biologice ca de exemplu:
variatia ionilor de Na+ si K+ in neuroni genereaza un impuls nervos (potential electric de actiune);
cresterea concentratiei ionilor de sodiu extracelular faciliteaza transportul unor aminoacizi in celule;
cresterea concentratiei ionilor de calciu in citoplasma initiaza contractia musculara sau secretia de enzime in celulele exocrine.
Functionarea acestor proteine canal determina modificari importante a gradientelor ionice si a potentialului electric, modificari care vor fi refacute rapid prin alte mecanisme de transport care necesita insa energie.
2.Transportul activ
Transportul activ prin membranele celulare se realizeaza cu ajutorul unor proteinenzime specifice membranelor biologice, care folosesc in acest transport fie energia rezultata din hidroliza ATP-ului, fie energia gradientelor chimice.
In ceea ce priveste mecanismul de transport al substantelor se cunosc de asemenea 2 tipuri de transport, dupa cum urmeaza:
transport activ primar, in cadrul caruia sunt cuprinse pompele ionice;
transport activ secundar, care se realizeaza cu ajutorul proteinelor transportor si prin care transportul unei specii moleculare, in sens contrar gradientului electrochimic, are loc pe baza energiei funizate de transportul simultan al altei specii in sensul gradientului electrochimic. In cadrul acestui tip de transport sunt incluse urmatoarele variante:
simport sau co-transport (transportul ambelor specii in acelasi sens): ;
antiport sau contra-transport (transportul ambelor specii in sens opus).
Descrierea pompelor ionice
|
1. Pompa de sodiu (Na+) si potasiu (K+) sau Na+, K+-ATP-aza
Plasmalema tuturor celulelor este polarizata, adica prezinta un potential de membrana cu valori cuprinse intre -20mV si -200mV. Fata citoplasmatica a plasmalemei este incarcata negativ, iar cea externa pozitiv. Exista doua proteine care joaca un rol foarte important in generarea si mentinerea potentialului de membrana: pompa de Na+ si K+, pe de o parte, si canalul de pierdere al K+, pe de alta parte. Concentratia intracelulara de K+, de aproximativ 400mM, este mult mai mare ca cea extracelulara (20mM). Pentru Na+ situatia este inversa, concentratia intracelulara fiind mult mai mica (50mM) fata de cea extracelulara (440mM). Proteina cu canalul de K+ permite difuziunea pasiva a K+ din celula la exterior (in sensul gradientului de concentratie). Dar prin aceasta interiorul celulei devine mai electronegativ si de aceea la o anumita valoare a potentialului de membrana (-75mV) tendinta K+ de a parasi celula datorita gradientului de concentratie este contrabalansata de tendinta K+ de a intra in celula datorita potentialului membranar. Deci gradientul electrochimic al K+ se anuleaza si numarul de ioni de K+ care ies din celula devine egal cu numarul celor care intra in celula. Dar canalul de K+ este usor permeabil si pentru Na+. Deci unii ioni de Na+ intra in celula conform gradientului electrochimic al Na+. Prin transport de sarcini pozitive la interiorul celulei scade potentialul membranar, ceea ce permite iesirea altor ioni de K+, iar prin repetarea proceselor descrise anterior s-ar ajunge la egalarea concentratiilor de Na+ si K+ de o parte si cealalta a membranei si la disparitia potentialului membranar, daca nu ar exista pompa de Na+ si K+ care pompeaza Na+ in afara celulei si K+ in citoplasma (sistem antiport), cu ajutorul energiei furnizate din hidroliza ATP-ului (Fig. 6).
Mediu intracelular Mediu extracelular
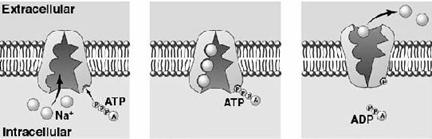
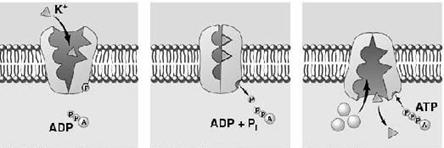
Fig. 6. Pompa ionica pentru ionii de sodiu si potasiu
Dupa 1957 s-a descoperit ca pompa de Na+ si K+ este aceeasi proteina cu Na+-K+-ATP-aza, o enzima care scindeaza ATP-ul in ADP si fosfat anorganic. Pentru fiecare molecula de ATP hidrolizata se pompeaza la exterior trei ioni de Na+ si la interior doi ioni de K+. Na+, K+ -ATP-aza poate scinda 100 de molecule de ATP pe secunda. De aceea, peste o treime din necesarul de energie al celulei este consumat de aceasta pompa, iar in celulele nervoase se ajunge ca pana la 70% din consumul energetic sa revina pompei. Prin pomparea la exterior trei ioni de Na+ si introducand in celula doi ioni de K+, proteina asigura potentialul de membrana, eliminand in afara celulei sarcini pozitive in exces. Mecanismul de functionare al acestei pompe este de tip ping - pong.
2. Pompa de calciu (Ca2+- ATP-aza)
Pompa de Ca2+ este importanta in mentinerea concentratiei scazute a ionului respectiv in citoplasma (10-7 M) fata de o concentratie mult mai mare a acestuia in mediul extracelular (10-3M). Gradientul mare de concentratie al Ca2+ asigura conditii pentru transmiterea de semnale de la exterior la interiorul celulei prin plasmalema. Reglarea concentratiei de Ca2+ in citosol este importanta si pentru procesele de secretie celulara si motilitate. Ca++-ATP-aza poate hidroliza 10 molecule de ATP intr-o secunda si pentru fiecare molecula hidrolizata pompeaza 2 Ca++ in afara citosolului (in mediul extracelular sau in canalele reticulului endoplasmatic) - Fig. 7.
Exterior
ADP + P ATP Loc de legare a calciului![]()
![]()
![]()
Loc de legare a ATP Citosol
![]()
Fig. 7. Pompa ionica pentru ionii de calciu (Mariana Sincai, 2000)
O caracteristica generala a pompelor ionice este reversibilitatea lor. Prin hidroliza ATP-ului ele mentin gradientele ionice, dar cand sunt puse in conditii care induc gradiente de concentratie ionica extrem de mari (depasind capacitatea pompelor); proteinenzimele pompelor ionice sintetizeaza ATP din ADP si fosfat anorganic.
3. Pompele protonice
In anumite tipuri de celule se remarca transportul ionilor prin membrana concomitent cu un transport de protoni (H+), mecanism dependent de ATP. Acest fenomen este remarcat in cazul lizozomilor sau a altor vacuole care au un pH acid.
In lizozomii celulelor animale se remarca existenta unui pH de 4,5 - 5,0, in timp ce pH-ul citosolului este de 7. Mentinerea acestui gradient protonic se realizeaza prin H+-ATP-aza. De asemenea, in membrana celulelor epiteliale ale vezicii urinare a fost identificata o H+-ATP-aza care acidifica urina.
Bacteria Halobacterium halobium are intr-o zona specializata a membranei sale, numita membrana purpurie, proteina bacteriorodopsina care functioneaza la lumina ca o pompa de protoni, expulzand protonii la exterior, creand astfel potentialul membranar.
O alta clasa, considerata a fi a patra clasa de proteinenzime transportoare dependente de hidroliza ATP-ului, a carui mecanism este inca insuficient cunoscut este considerata a fi clasa glicoproteinelor de membrana care prin hidroliza ATP-ului transporta din celula in mediul extracelular o gama larga de medicamente sau droguri utilizate in tratamentul tumorilor (vinblastina, colchicina etc.). Majoritatea acestor substante sunt hidrofobe si patrund rapid in celula.
Transportul activ care foloseste energia gradientelor ionice generata prin hidroliza de ATP
Acest mecanism consta in faptul ca energia rezultata din hidroliza ATP-ului este utilizata pentru a genera un gradient crescut al Na+ extracelular, gradient care este utilizat apoi ca sursa de energie pentru patrunderea glucozei si aminoacizilor in celule.
De importanta foarte mare este transportul glucozei prin plasmalema celulelor din mucoasa intestinala si a celulelor din tubii renali prezinta importanta foarte mare. Glucoza este transportata in celule impotriva gradientului de concentratie cu ajutorul unei proteine transportoare care leaga si Na+, prin system simport. Cu cat diferenta dintre concentratia intracelulara si cea extracelulara a Na+ este mai mare, cu atat mai mare este si viteza de transport a glucozei. Na+ care intra in celula cu glucoza este pompat in afara de catre Na+K+-ATP-aza (Fig. 8). Transportul aminoacizilor se realizeaza tot prin sisteme simport cu Na+, existand cel putin cinci proteine diferite in plasmalema celulelor animale, una pentru fiecare grup de aminoacizi inruditi structural.
In membrana bacteriilor, cele mai multe sisteme de transport folosesc in transportul glucidelor si aminoacizilor ca ion simportat -H+.
Translocarea de grup se intalneste la unele bacterii si consta in procesul de fosforilare a unor glucide in cursul trecerii lor prin plasmalema. Sistemul de fosforilare (fosfoenolpiruvat transferaza) este complex, implicand patru proteine si fosfoenolpiruvatul ca sursa de energie.Prin fosforilare, glucidele devin ionizate si nu mai pot parasi celula. De asemenea, fosforilarea glucidelor mentine o concentratie scazuta de glucide libere in celula, ceea ce permite, conform gradientului de concentratie, o captare continua a acestora in celula.
Tot la bacterii se mai intalnesc sisteme pentru transportul activ al glucidelor si aminoacizilor care implica proteine hidrosolubile localizate in spatiul dintre plasmalema si peretele celular.
Exterior
Transport Gradient de
concentratie glucoza Na
![]()
![]()
Fig. 8. Transportul simport al glucozei cu ionii de sodiu (Mariana Sincai, 2000)
Particularitati ale sistemelor de transport activ prin membrane si importanta acestora
Sistemele simport, in care patrunderea unei substante in celule, in mod obisnuit a Na+, spre gradientul de concentratie mai scazut impreuna cu o alta substanta impotriva gradientului de concentratie al acesteia, cum este glucoza, sunt deosebit de importante pentru metabolismul celular. Insa, la fel de importante pentru celula sunt si sistemele antiport, in care patrunderea unor ioni este cuplata cu iesirea altora.
Un astfel de sistem antiport bine studiat este cel intalnit in celulele musculare cardiace si in alte celule animale in care se reduce concentratia interna de Ca2+ prin patrunderea concomitenta a doi sau trei ioni de Na+.
Na+-K+-ATP-aza din membrana celulelor musculare cardiace creeaza gradientul de concentratie al ionilor de Na+ (mare la exteriorul celulei) care este folosit ca energie (forta) pentru exportul ionilor de Ca2+. Astfel, medicamente cum sunt: digoxin (digitalina) si ouabain, au o mare insemnatate clinica, ele crescand forta de contractie a muschiului cardiac prin inhibarea Na+-K+-ATP-azei (Fig. 9). Ca urmare, in celulele cardiace se mentine un nivel mai ridicat al Na+ si sistemul antiport Na+/Ca2+ functioneaza mai putin eficient la un gradient mai scazut al ionilor de Na+. Ca urmare mai putini ioni de Ca2+ sunt exportati, nivelul de Ca2+ intracelular mentinandu-se crescut, astfel ca muschiul cardiac se poate contracta mai des si mai puternic.
Na+K+-ATP-aza Sistem antiport Na+/Ca+ Digoxin 
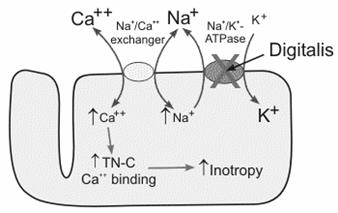
Fig. 9. Sistemul antiport Na+/Ca2+ din celula musculara cardiaca
Un alt sistem antiport important, existent in membrana eritrocitelor este cel implicat in schimbul anionilor, respectiv al ionilor negativi de Cl- si HCO¯3. Acest fenomen este posibil deoarece permeabilitatea membranei eritrocitelor este de 100000 de ori mai mare decat a altor celule.
Dioxidul de carbon eliberat de celulele tesuturilor difuzeaza in capilarele sanguine si patrunde in eritrocite. In forma sa gazoasa, dioxidul de carbon se dizolva slab in solutii apoase cum este plasma sanguina, dar in prezenta anhidrazei carbonice, produsul rezultat disociaza in H+ si HCO¯3. Procesul are loc in timp ce oxigenul se desprinde de hemoglobina, in locul lui fiind legat H+. Anionul HCO¯3 este scos din eritrocite concomitent cu patrunderea ionilor de clor, printr-un sistem antiport realizat de o proteina transmembranara. Daca acest schimb de anioni nu ar avea loc, HCO¯3 s-ar acumula in eritrocite pana la un nivel toxic, pe masura ce este generat CO2, citoplasma eritrocitelor devenind alcalina. Sistemul antiport HCO¯3 - Cl- este cel care asigura mentinerea pH-ului eritrocitelor la un prag neutru.
La pulmoni, HCO¯3 difuzeaza in eritrocite, prin schimb cu anionii de Cl-, unde se combina cu ionii de H+ inlocuiti in hemoglobina de catre O2. H2C03 in prezenta anhidrazei carbonice se desface in H20 si CO2, care difuzeaza in afara eritrocitelor unde concentratia lui este mai mica. De specificat ca aceasta proteina de membrana nu permite curgerea unidirectionala a anionilor dintr-o parte in alta a membranei, decat printr-un schimb echilibrat in sens antiport.
Aceste sisteme antiport care regleaza pH-ul in celulele animale sunt foarte importante, deoarece pentru a creste si a se multiplica, pH-ul citoplasmei trebuie sa se mentina intre limite relativ stranse, respectiv 7,2 - 7,4.
Anhidraza carbonica Celulele tesuturilor
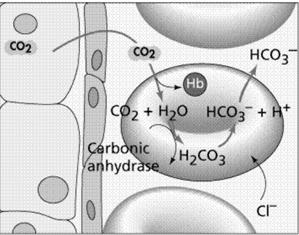
Fig. 10. Sistemul antiport Cl- si HCO¯3 din membrana eritrocitelor aflate la nivel tisular
Alveola pulmonara Anhidraza carbonica
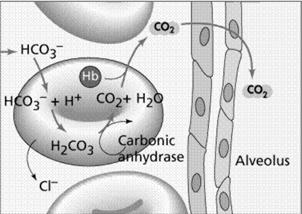
Fig. 11. Sistemul antiport Cl- si HCO¯3 din membrana eritrocitelor aflate la nivel pulmonar
Un sistem antiport important este semnalat in membrana apicala a celulelor parietale din epiteliul mucoasei stomacului, celule care secreta acidul clorhidric, parte componenta a sucului gastric. Celulele parietale (acidofile) contin H+K+-ATP-aza care scoate din celula doi protoni de H+ si introduce un ion de K+. Iesirea H+ este insotita de un transport pasiv de Cl-, formandu-se, in lumenul gastric, acidul clorhidric. Prin iesirea H+, in celula creste concentratia de OH-. Pentru a asigura pH-ul neutru, excesul de OH- se combina cu CO2 formand HCO¯3, anion care este transportat extracelular prin membrana laterobazala, concomitent cu patrunderea Cl-, prin acelasi sistem luat in discutie.
.2. Transportul substantELOR prin membrana CELULARa CU AJuTORUL VEZICULELOR
Un mod special de transport prin membranele biologice il reprezinta transportul cu ajutorul veziculelor. Se disting trei procese: exocitoza, endocitoza si transcitoza, dupa cum transportul are loc din citoplasma la exterior, spre interiorul celulei, sau, respectiv, prin vezicule care strabat complet celula dintr-o parte in cealalta.
Exocitoza
Este procesul prin care vezicule din citoplasma, care contin diverse substante, fuzioneaza cu plasmalema si sunt eliminate in afara celulei (Fig. 12).
Vezicula de transport Citoplasma Mediu extracelular Plasmalema Substante transportate
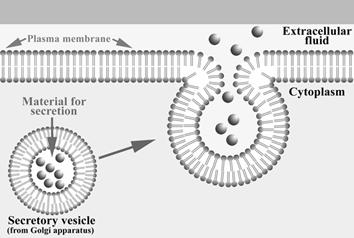
Fig. 12. Exocitoza
In organismele multicelulare exista doua tipuri de exocitoza, respectiv:
- constitutiva, independenta de ionii de Ca2+;
- neconstitutiva, dependenta de ionii de Ca2+.
Exocitoza constitutiva are loc in toate celulele si serveste fie la eliberarea unor substante rezultate in urma digestiei in afara celulei, fie la livrarea catre membrana celulara a unor proteine nou-sintetizate (produsi de secretie din celulele glandulare).
Exocitoza care are loc la nivelul sinapselor chimice neuronale este dependenta de ionii de Ca2+ si deserveste semnalizarea interneuronala (in cazul sinapsei neuronale, impulsul electric e convertit in informatie chimica, prin intermediul exocitozei).
Etapele exocitozei cuprind: transportul veziculelor, capturarea veziculelor langa membrana tinta, ancorarea acestora la membrana, conditionarea si fuzinarea.
Acest fenomen prezinta o importanta deosebita pentru celule, fiind necesar in:
secretia enzimelor, hormonilor si anticorpilor din celule;
mentinerea si reinnoirea membranei celulare;
eliberarea de neurotransmitatori din presinapsele neuronale;
transportul proteinelor membranare integrale;
reactia acrozomala in timpul fertilizarii ovulului;
prezentarea anticorpilor in timpul raspunsului imun;
reciclarea receptorilor membranari;
comunicarea dintre celule;
Endocitoza
Reprezinta inglobarea in celule a unor substante cu ajutorul veziculelor.
Dupa natura substantelor care patrund in celula, exista doua tipuri de endocitoza, respectiv:
fagocitoza, prin care se inglobeaza particule solide(gr. fagein= a manca);
pinocitoza, prin care celula inglobeaza macromolecule in solutie (gr. pinein= a bea).
In ambele cazuri de endocitoza, substantele patrund in celula inglobate in vezicule desprinse din plasmalema, vezicule numite endozomi, respectiv fagozomi sau pinozomi.
Particulele solide mari sunt inglobate numai de celule specializate numite fagocite, in timp ce moleculele in solutie sunt preluate de majoritatea celulelor.
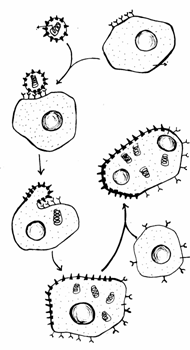
Fagocitoza este o modalitate de hranire a protozoarelor. La vertebrate insa, fagocitoza detine un rol deosebit de important in procesele de aparare ale organismului, pe aceasta cale fiind inglobate si distruse bacteriile, parazitii, substantele straine, celulele imbatranite, celulele degenerate sau distruse, celulele tumorale maligne, etc.
La mamifere si la om fagocitele sunt de doua tipuri: macrofage si neutrofile, ambele avand originea in celulele stem ale maduvei hematogene osoase (Fig. 13).
Membrana celulara Fagozom
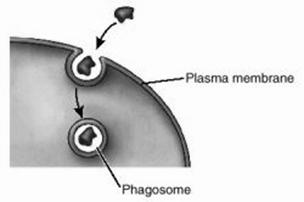
Fig. 13. Fagocitoza
Procesul de fagocitoza cuprinde mai multe faze:
chemotactismul, respectiv atragerea si deplasarea fagocitelor spre componentele straine organismului (bacterii, toxine, resturi celulare) care emit semnale chemotactice;
recunoasterea si atasarea fagocitelor de particulele straine prin receptorii specifici care recunosc anumiti mesageri (liganzi);
inglobarea de catre fagocit a particulei straine intr-o vezicula internalizata in citoplasma numita fagozom si care ulterior va fi supusa actiunii lizozomilor.
O mare parte din substantele rezultate in urma digestiei de catre lizozomi vor trece in citoplasma si vor fi utilizate in diferite reactii metabolice. Substantele nedigerate sau neutilizabile sunt eliminate din celula prin exocitoza.
Pinocitoza este de doua tipuri:
pinocitoza independenta de receptori;
pinocitoza dependenta sau mediata de receptori.
Pinocitoza independenta de receptori se mai numeste si endocitoza in faza fluida si este intalnita la numeroase celule. Prin acest proces se introduc in celula mari cantitati de macromolecule in solutie, insa volumul celulei se mentine constant deoarece simultan cu endocitoza are loc si un fenomen de exocitoza prin care se asigura totodata constanta de suprafata a plasmalemei. In acest fel, prin endocitoza si exocitoza, componentele plasmalemei sunt recirculate si reciclate.
Pinocitoza dependenta de receptori se realizeaza cu ajutorul receptorilor din plasmalema care recunosc anumite macromolecule specifice (Fig. 14).
Trecerea in citoplasma a
substantelor utile Lizozom secundar Digestie Vacuola cu LDL Lizozom primar Endocitoza mediata de receptori Colesterol Lipide Proteine Receptor Particula de LDL Reciclarea membranelor
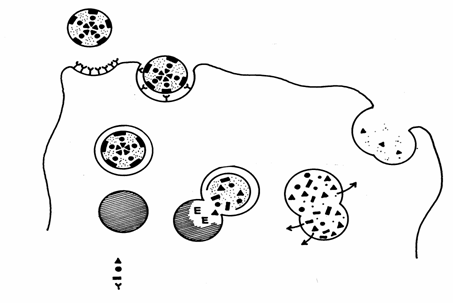
Fig. 14. Endocitoza lipidelor cu mica densitate (Mariana Sincai, 2000)
Complexul ligand - receptor determina aparitia unor depresiuni ale plasmalemei numite caveole (coated pits), care apoi devin vezicule de endocitoza. In aceste vezicule, ca urmare a concentrarii anumitor liganzi, concentratia substantelor este diferita fata de cea din citosol.
Un fenomen de endocitoza mediata de receptori, foarte important pentru organism, este captarea colesterolului in celulele animale. Majoritatea colesterolului este transportat in sange sub forma complexelor LDL sau lipoproteine cu densitate mica (low density lipoproteins). Acestea sunt particule sferice de 22 nm care au la interior 1500 molecule de colesterol esterificat cu acizi grasi, iar la exterior prezinta un stat dublu lipidic asociat cu o proteina.
O celula animala care necesita colesterol produce receptori LDL, dupa care complexele LDL-receptor sunt introduse prin endocitoza in celula. Prin fuzionarea acestor vezicule cu lizozomii, esterii de colesterol sunt hidrolizati, iar colesterolul liber trece in citosol, inhiband formarea receptorilor pentru LDL si sinteza colesterolului.
In anumite boli genetice se produc receptori LDL anormali (care nu pot lega LDL sau nu se pot deplasa in caveole) si ca urmare LDL nu se poate capta in celula, aceasta sintetizand colesterolul in cantitati foarte mari, producandu-se o hipercolesterolemie cu consecinte mai ales de arteroscleroza.
Transcitoza este un fenomen de transport prin vezicule care se deplaseaza de la o suprafata la alta a celulei (Fig. 15). Este o forma de transport a macromoleculelor prin celulele endoteliului vascular. Astfel, se formeaza vezicule care se deplaseaza in celula dinspre fata luminala spre fata interstitiala sau invers, realizandu-se schimburi de substante intre plasma sanguina si lichidul intercelular. Uneori cand viteza de formare si de circulatie a veziculelor este mare, ele pot fuziona si forma canale care strabat celula endoteliala de pe o fata pe alta.
Lumen vascular
Matrice extracelulara
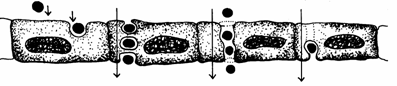
Fig. 15. Transcitoza prin celulele endoteliale (Mariana Sincai, 2000)
Particularitati privind transportul substantelor prin endocitoza
Prin acest fenomen de endocitoza mediata de receptori patrund in celule numeroase virusuri si toxine.
In ceea ce priveste virusurile care traiesc la animale, multe tipuri sunt alcatuite dintr-un material genetic viral si o capsida proteica peste care se gaseste o membrana externa alcatuita dintr-un bistrat de fosfolipide. In mod obisnuit, virusurile se formeaza prin inmugurirea plasmalemei, invelisul fosfolipidic al virusurilor derivand astfel din membrana celulara. Membrana virala contine de asemenea una sau mai multe glicoproteine specifice virusului, fara de care acesta nu ar putea infecta celula tinta. Particulele virale sunt endocitate in vezicule cu pH scazut, dupa care membrana virala fuzioneaza cu membrana veziculei, astfel incat acidul nucleic viral este eliberat in citosol, initiindu-se replicarea acestuia. Cresterea pH-ului in endozomi inhiba infectia virala.
In cazul unor virusuri ca: paramixovirusurile, virusul Sendai si virusul HIV (virusul imunodeficientei sau AIDS), invelisul acestora fuzioneaza direct cu plasmalema celulelor fara o endocitoza prealabila (Fig. 16).
Astfel, limfocitele T, macrofagele si unele celule ale creierului prezinta in membrana lor receptori pentru virusul HIV, respectiv o glicoproteina numita CD4.
HIV infecteaza si omoara numai aceste celule, insa moartea este cauzata de o blocare completa a intregului sistem imun. Infectia virala se raspandeste direct de la o celula la alta fara interventia unui virus liber. In limfocitele T, odata cu replicarea virusului se produce si o mare cantitate de glicoproteina virala care, in parte, intra in structura unor noi virusuri iar o alta parte se leaga de proteina CD4 a limfocitelor T invecinate si neinfectate. Ca urmare are loc o fuziune a celulelor infectate cu celule neinfectate rezultand sincitii multinucleate.
Limfocit T neinfectat