|






 |  |

| Biologie | Botanica | Chimie | Didactica | Fizica | Geografie |
| Gradinita | Literatura | Matematica |
Biologie
|
|
Qdidactic » didactica & scoala » biologie Variabilitatea numarului de cromozomi - tipuri de mutatii ale numǍrului de cromozomi |
Variabilitatea numarului de cromozomi - tipuri de mutatii ale numǍrului de cromozomi
VARIABILITATEA NUMARULUI DE CROMOZOMI
Numarul, forma si marimea cromozomilor sunt trasaturi caracteristice stabile pentru diferite specii. Acestea alcatuiesc cariotipul, care este un criteriu de recunoastere.
In celule reproducatoare este prezent un singur set cromozomal (starea haploida, notata simbolic cu n).
Numarul de baza, numarul monoploid, sau setul haploid al unei specii diploide, se noteaza cu x.
Schimbarea numarului de cromozomi, poate avea loc spontan, sau artificial sub actiunea factorilor mutageni. Modificarea numarului de cromozomi, constiutuie una din caile evolutiei speciilor, fiind corelata cu modificarea cantitatii de ADN din celule.
Mutatiile cromozomale numerice sunt cunoscute si sub denumirea de mutatii de genom si sunt foarte numeroase la plante.
1. TIPURI DE MUTATII ALE NUMǍRULUI DE CROMOZOMI
Mutatiile la nivelul numarului de cromozomi, sunt clasificate in functie de prezenta sau absenta unor cromozomi particulari sau a unor seturi intregi de cromozomi.
1. 1. EUPLOIDIA
Euploidia este reprezentata de starea celulara in care numarul de cromozomi corespunde cu numarul de baza sau cu un multiplu al acestuia. Indivizii afectati de euploidie se numesc euploizi.
Dupa numarul de seturi cromozomale, euploidia poate fi:
monoploidie si haploidie, atunci cand numarul de cromozomi din celulele somatice este egal cu numarul de baza (x), respectiv cu cel gametic (n);
poliploidie, numarul de cromozomi din celulele somatice cuprind mai mult decat doua genomuri, fiind triploizi 3x, tetraploizi 4x, pentaploizi 5x, hexaploizi 6x etc.
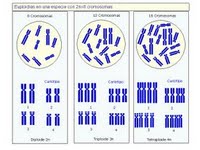
In aneuploidie modificarea numarului de cromozomi este restransa la mai putin de un set cromozomal si afecteaza numai anumiti cromozomi, prin adaugarea a unu, doi cromozomi si chiar mai multi (polisomie) cromozomi, sau pierderea unor cromozomi (oligosomie).
Prezentarea schematica a diferitelor schimbari in numarul de cromozomi este ilustrata in fig. 1.
Poliploidia este fenomenul de multiplicare a numarului cromozomal de baza. Autoploidia rezulta in urna multiplicarii setului cromozomal de baza propriu, sau in urma hibridarii unor indivizi poliploizi care apartin la aceiasi specie. La autoploizi, seturile cromozomale sunt omoloage.
Fig. 1. Schimbari in numarul de cromozomi
![]() 1. Euploidia
1. Euploidia
- monoploidia
- haploidia
- poliploidia
![]() - artioploidia
- artioploidia
![]() - autoploidia - perisoploidia
- autoploidia - perisoploidia
- poliploidia
![]() -
genomala
-
genomala
- aloploidia - segmentala
2. Aneuploidia
![]()
- monosomia
- oligosomia
- nulisomia
![]()
- trisomia
- polisomia
- tetrasomia
Acestia pot fi: artioploizi (autoploizi, cu numarul de baza multiplicat intr-un numar impar: 3x, 5x, 7x etc).
Multiplicarea numarului de cromozomi atrage dupa sine unele modificari ereditare, care vor fi protejate si favorizate de selectie daca sunt utile speciei.
1. 1. 1. Caracteristicile autoploizilor
In celulele poliploide apar o serie de modificari care sunt identificate prin investigare microscopica. Datorita numarului sporit de cromozomi, celulele sunt mai mari, diametrul celulelor somatice este mai mare si contin un numar sporit de cloroplaste. In celula poliploida numarul cromocentrilor nucleolari este de asemenea mai mare. Diametrul graunciorilor de polen de la organismele poliploide, ca si numarul porilor germinativi a polenului este mai mare.
Macroscopic se disting mai multe particularitati, habitusul plantelor este mai mare si mai viguros, cu numar mic de tulpini, lastari sau ramificatii, dar de dimensiuni mult mai mari. Plantele au flori mai putine dar mari si intens colorate. Aparatul foliar este alcatuit dintr-un numar mai mic de frunze, de dimensiuni mai mari si de un verde intens.
La plante, poliploidia este corelata si cu importante modificari fiziologice si biochimice. Astfel, plantele prezinta o cantitate de clorofila si o capacitate sporita de fotosinteza, rezistenta crescuta la stresul climatic si agenti patogeni, manifestand, in general, o buna homeostazie (adaptabilitate) la conditii de mediu nefavorabile, asociate insa cu tardivitate.
Ca urmare a efectului de dozaj, plantele poliploide au o capacitate crescuta de sinteza a substantelor proteice, a glucidelor, a vitaminelor, pigmentilor etc.
Fertilitatea autoploizilor, in general, este mai scazuta, mai ales la perisoploizi, deoarece apar diferite anomalii in desfasurarea meiozei. Anomaliile sunt frecvente in profaza I, deoarece alaturi de bivalenti, se formeaza si uni-, tri- si tetra- valenti. Multivalentii segrega dezordonat (nebalansat) in anafaza I ceea ce duce la formarea de gameti neechilibrati din acest considerent se recomanda utilizarea plantelor poliploide numai atunci cand din punct de vedere economic, intereseaza masa vegetativa si nu productia de seminte (plante furajere, plante ornamentale, sfecla de zahar etc.).
Segregarea, in cazul autoploizilor este mult mai complicata decat in mod normal. Daca o forma diploida heterozigota (Aa) produce doua tipuri de gameti (A) si (a), in F2 segregarea genotipica este 1/4 AA + 2/4 Aa + 1/4 aa, iar fenotipica 3/4 dominanti : 1/4 recesivi. In descendenta autogama a unui autotetraploid AAaa, pot exista pentru acelasi locus 5 genotipuri: forme homozigote AAAA (quadruplex) si aaaa (multiplex) respectiv, forme heterozigote AAAa (triplex), AAaa (duplex), Aaaa (simplex).
In consecinta, numarul tipurilor de gameti si a claselor de segregare genotipica si fenotipica este mult mai mare.
Raportul de segregare la autotetraploizi in cazul in care alela dominanta este prezenta intr-o singura doza este prezentat in tabelul 1.
Tabelul 1.
Raportul de segregare al autotetraploizilor cu o singura alela dominanta
(dupa R.C. Williams, 1985)
|
Parinti |
Raportul gametic |
Raportul fenotipic |
||||
|
A1A2a3a4 x A1A2a3a4 |
1 AA : 4 Aa :1 aa |
35 A : 1 a |
||||
|
A1A2a3a4 x A1a2a3a4 |
1 AA : 4 Aa :1 aa 1 Aa : 1 aa |
11 AA : 1 a |
||||
|
A1A2a3a4 x a1a2a3a4 |
1 AA : 4 Aa :1 aa toti sunt aa |
5 A : 1 a |
||||
|
A1a2a3a4 x A1a2a3a4 |
1 Aa : 1 aa |
3 A : 1 a |
||||
|
A1a2a3a4 x a1a2a3a4 |
1 Aa : 1 aa toti sunt aa
|
1 A : 1 a |
Dupa mecanismul de producere, autoploizii pot fi mitotici si meiotici.
Autoploizii mitotici apar ca urmare a deranjarii diviziunilor mitotice normale. Sub actiunea unor factori externi este posibil ca uneori sa nu se formeze fusul de diviziune si astfel cromozomii separati in cromatide, nu mai migreaza in anafaza spre polii celulei si ca urmare membrana nucleara, va ingloba un numar dublu de crmozomi, rezultand un tetraploid.
Poliploizi mitotici mai pot apare si atunci cand se formeaza un nucleu de restitutie, adica se produce diviziunea normala a nucleului, fara ca diviziunea citoplasmei sa aibe loc.
Autoploizii meiotici rezulta datorita alterarii procesului normal al meiozei. In aceasta situatie, se poate intampla ca in profaza I cromozomii sa nu formeze bivalenti sau sa se formeze un nucleu de restitutie (prin migrarea in anafza I la un singur pol a tuturor cromozomilor). Acest fapt, determina formarea unor gameti diploizi, care prin fecundare vor da nastere unui autotetraploid.
1. 1. 2. Inducerea artificiala a poliploidiei
Pentru crearea de poliploizi artificiali s-au pus la punct numeroase metode de modificare a diviziunii celulare.
Se preteaza la poliploidizare acele specii care prezinta un numar mic de cromozomi, la care in primul rand se utilizeaza masa vegetativa si mai putin semintele, precum si speciile care prezinta un grad ridicat de sterilitate, creandu-se posibilitatea de a se multiplica vegetativ.
Cele mai utilizate metode de inducere a poliploidiei sunt:
tratamente cu socuri de temperatura, ridicate sau scazute, aplicate la diferite organe, tesuturi sau celule aflate in diviziune. Socurile de temperatura deregleaza aparatul mitotic (fusul nuclear) generand artioploidie;
metoda centrifugarii si a iradierii, conduc la inhibarea fusului de diviziune, microtubulii nu se asambleaza ca sa formeze firele acromatice sau sunt distrusi, fiind posibila aparitia unor nuclei de restitutie;
metoda regenerarii se bazeaza pe dublarea numarului de cromozomi prin endomitoza, frecvent intalnita in culturile de celule si calusurile de cicatrizare (punctul de altoire), celule care se multiplica in prezenta heteroauxinei;
metoda poliembrioniei se bazeaza pe predispozitia unor specii (Zea mays, Secale cereale etc.), de a forma seminte cu doi sau mai multi embrioni, dintre care unii pot fi poliploizi;
metoda tratamentului chimic, cu colchicina sau cu proto-oxid de azot, este cea mai eficienta si mai frecvent utilizata.
![]() Metoda a fost elaborata
de A.F.
Blakeslee si O.T. Avery (1937) si se
bazeaza pe actiunea colchicinei asupra fusului acromatic din celule
in diviziune. Colchicina este un alcaloid extras din bulbi de
brandusa de toamna (Colchicum
autumnale) care datorita actiunii ei citostatice, are capacitatea
de a bloca formarea fusului si astfel, dupa o mitoza, celulele
vor fi artioploide (2n = 4n) dupa doua mitoze celulele
vor avea 8n cromozomi
Metoda a fost elaborata
de A.F.
Blakeslee si O.T. Avery (1937) si se
bazeaza pe actiunea colchicinei asupra fusului acromatic din celule
in diviziune. Colchicina este un alcaloid extras din bulbi de
brandusa de toamna (Colchicum
autumnale) care datorita actiunii ei citostatice, are capacitatea
de a bloca formarea fusului si astfel, dupa o mitoza, celulele
vor fi artioploide (2n = 4n) dupa doua mitoze celulele
vor avea 8n cromozomi
![]()
![]() (2n 4n 8n).
(2n 4n 8n).
Solutia apoasa de colchicina in concentratie de 0,1 - 1,0% poate fi utilizata pentru tratamentul semintelor, a varfurilor vegetative si radacinilor plantelor, a mugurilor, a florilor sau inflorescentelor.
Sub influenta colchicinei, diviziunea sufera unele modificari, fiind denumita C-mitoza si C-meioza. Colchicina, nu are actiune semnificativa asupra cromozomilor, clivarea centromerului si separarea cromatidelor este normala, in interfaza dintre diviziuni formandu-se cromozomi cu morfologie specifica. Colchicina blocheaza polimerizarea tubulinei si in acest fel, formarea fusului de diviziune este anihilata.
In urma colchicinizarii, in mitoza apar schimbari specifice si anume: profaza decurge normal. In metafaza, in absenta fusului de diviziune, cromozomii nu se dispun la centrul celulei, pentru a forma placa metafazica, ramanand dispersati in citoplasma (pseudometafaza sau C-metafaza). La cromozomii puternic condensati, cromatidele apar desfacute, fiind unite numai la centromer (configuratia literei X, cromozomi-X sau cromozomi-C). Cele patru cromatide ale unei perechi de cromozomi raman in aceeasi celula, formand nucelul de restitutie cu un numar dublu de cromozomi.
Meioza celulelor colchicinizate se caracterizeaza prin absenta fenomenelor sinaptice. Astfel ca in C-metafaza, cromozomii nu formeaza bivalenti, ei raman ca univalenti. In C-anafaza I, cromatidele surori nu se separa, toti cromozomii fiind inclusi intr-un singur nucleu, denumit nucleu de restitutie, care va avea un numar dublu de cromozomi. Meioza de acest tip, genereaza formarea gametilor diploizi, care in urma fecundarii, vor da nastere zigotilor tetraploizi.
Din semintele tratate cu colchicina, pot aparea plante noi, in totalitate poliploide. Aplicarea tratamentului la parti vegetative ale plantei, determina formarea de himere, tesuturi mixoploide, in care alterneaza celule diploide si poliploide.
1. 1. 3. Importanta autoploidiei
Poliploidia, este un proces cu importanta deosebita, atat pentru evolutie cat si pentru ameliorarea plantelor.
In evolutie, autoploidia reprezinta o importanta sursa de variabilitate, numeroase specii avand o origine poliploida (cartoful 2n = 48; alunele de pamant 2n = 40; cafeaua 2n = 22, 44,66, 88 etc.)
A. Müntzing (1967), considera ca exista o corelatie pozitiva intre numarul de cromozomi si durata vietii unei plante. Numeroase specii perene au provenit din plante anuale cu un numai mic de cromozomi. Astfel, Sorghum sudanensis, forma anuala, are n = 10 cromozomi in timp ce forma perena, Sorghum helepense are n = 20 cromozomi; Helianthus annus, cu n = 17, este o specie anuala, pe cand Helianthus tuberosus, specie perena are n = 51 cromozomi.
Dupa studiile lui G. Tischler (1967) cu privire la repartitia pe glob a acestui fenomen se constata ca frecventa poliploidiei creste spre poli si scade spre zonele calde. In regiunile arctice, speciile poliploide reprezinta 92% din flora.
Pentru ameliorarea plantelor, autoploizii constituie un material biologic de o importanta deosebita.
Poliploidia determina sporirea cantitatii de ADN, implicit a numarului de gene si respectiv a variabilitatii genetice generate de efectul de doza.
La plante, poliploidia este corelata cu importante modificari morfologice, fiziologice si biochimice. In consecinta, in programele de ameliorare a plantelor sunt utilizate formele autoploide, valorificandu-se anumite particularitati, cum ar fi, vigoarea vegetativa, continut ridicat in proteine sau in vitamine, pigmentii, precum si rezintenta sporita la temperaturi excesive, seceta, boli si daunatori.
Din punct de vedere practic poliploidia are o importanta economica deosebita. Astfel, la triploizi, la care sterilitatea impiedica reproducerea sexuata, nu se formeaza seminte, ceea ce este deosebit de util in cazul strugurilor pentru stafide (3n = 57), pepenelui verde (3n = 33), bananierului (3n = 33) sau ananasului (3n = 75), forme preferate de consumatori.
La sfecla de zahar, triploizii au cel mai ridicat continut de zahar si cea mai mare productie de radacini.
Formele autoploide sunt importante pentru sectorul plantelor ornamentale, deoarece dimensiunile florilor sunt mai mari, pigmentatia este mult mai intensa si florile se pastreaza mai mult (lalele, crizanteme, garoafe, narcise).
Producerea de seminte este un alt domeniu in care autoploidia are o mare aplicabilitate. Datorita segregarii deficitare si incetinita, prin autoploidie se pot obtine combinatii hibride, cu heterozis mai stabil in succesiunea generatiilor.
1. 1. 4. Haploidia
Haploidia, este fenomenul prin care numarul de cromozomi din celulele somatice se reduce la jumatate. In celulele tesutului somatic, plantele haploide au numarul de cromozomi egal cu cel din celulele gametice.
Dupa modul in care se formeaza, formele haploide se grupeaza in doua categorii: monoploide si polihaploide.
Monoploidia reprezinta starea celulelor somatice, sau a organismelor, la care numarul de cromozomi corespunde cu numarul de baza, poseda un singur set cromozomal (x). Monoploizii, sunt haploizii speciilor diploide, posedand numarul gametic de cromozomi.
Polihaploidia reperezinta reducerea la jumatate a numarului de cromozomi din tesut somatic al plantelor poliploide. Polihaploizii, sunt haploizii speciilor poliploide.
1. 1. 5. Caracteristicile haploizilor
La nivelul structurii microscopice, haploidia se coreleaza cu reducerea volumului nuclear si a intregii celule. Celulele somatice sunt mai mici si inglobeaza un numar redus de cloroplaste, acestea fiind repartizate neuniform pe unitatea de suprafata a frunzei.
Toate caracteristicile morfologice ale haploizilor sunt subdimensionate, vigoarea este scazuta, talia este redusa, frunzele sunt mai putine, inguste si de culoare mai deschisa.
Reducerea taliei si a vitalitatii sunt mai pregnante la haploizii proveniti de la plante alogame. Existenta unui singur set cromozomal, permite manifestarea tuturor genelor recesive, atat a celor utile cat si a celor defavorabile. La plantele autogame consecintele morfologice si fiziologice nu sunt atat de acute, deoarece in starea diploida a organismului, homozigotia este o conditie normala.
Haploizii au o fertilitate extrem de redusa, la monoploizi sterilitatea este foarte ridicata deoarece meioza se desfasoara anormal. Prezenta in celulele mama a gametilor, a unui singur set cromozomal, impiedica formarea bivalentilor si repartizarea balansata a acestora in anafaza I a meiozei. Cromozomii se repartizeaza neregulat spre cei doi poli, producand nuclei anormali si implicit sterilitatea. Daca in anafaza I a meiozei, intregul set de cromozomi se deplaseaza la un singur pol al celulei, meioza homeotipica se desfasoara normal si ca rezultat se vor forma 50% spori normali si functionali. Probabilitatea ca migrarea sa aibe loc spre un singur pol al celulei este de (1/2)x-1, unde x este numarul cromozomi. Relatia asigura cunoasterea frecventei gametilor viabili, in functie de numarul cromozomilor. Datorita starii de hemizigotie (prezenta intr-un locus a unei singure alele), gametii monoploizilor sunt identici. Un grad ridicat de sterilitate se inregistreaza si la organismele polihaploide, provenite din indivizi cu grad ridicat de poliploidie, datorita formarii de univalenti, trivalenti etc. Prin formarea bivalentilor, materialul genetic se poate distribui echilibrat in gameti si astfel acestia vor fi functionali.
1. 1. 6. Posibilitati de obtinere a haploizilor
Dupa R. Ringer, citat de P. Raicu (1997), frecventa cu care apar haploizii in mod spontan este redusa. Haploidia este conditionata de genotip, astfel la Gossypium barbadense variaza in limitele 0,025-0,037%, la Datura stramonium este de 0,018%, la Zea mays, de 0,05%, la Anthirrhinum majus este 0,01%, la Triticum monococcum este de 0,05%, la Triticum aestivum de 0,025% etc.
Posibilitatile de obtinere a haploizilor sunt:
1. androgeneza, atunci cand fecundarea este simpla (embrionii se formeaza fara fecundare, o singura spermatie polinica fecundeaza nucleul secundar, din care rezulta endospermul).
2. ginogeneza, prin partenogeneza, rezulta embrioni din oosfera nefecundata si apogamie, atunci cand embrionul haploid se dezvolta din sinergide sau antipode. Haploizii se pot forma ca urmare a fenomenului de poliembrionie, adica aparitia de embrioni gemeni, din care unul poate fi haploid.
Un exemplu de organism haploid natural obtinut prin partenogeneza (ginogeneza) este trantorul, masculul speciei Apis melifera, cu n = 16 cromozomi (starea diploida este 2n = 32).
1. 1. 7. Inducerea artificiala a monoploidiei
Caile de producere a formelor monoploide pot fi in vivo cat si in vitro.
Metodele utilizate pentru inducerea in vivo a haploidiei sunt:
- iradierea polenului cu radiatii ionizante sau ultraviolete in vederea distrugerii unei spermatii, ce duce la formarea de haploizi dintr-o oosfera nefecundata;
- polenizarea intarziata si tratamentele cu socuri de temperatura imediat dupa polenizare;
- indepartarea staminelor florale, fapt ce impiedica polenizarea normala;
- inactivarea nucleilor spermatici prin tratamente cu diferite substante chimice, care favorizeza o dezvoltare partenogenetica;
- inhibarea cresterii tubului polinic prin tratament cu hidrazida maleica.
Metodele de inducere in vitro a haploidiei sunt:
culturi de antere, din graunciori de polen imaturi sau in culturi de ovule nefecundate, pe medii artificiale.
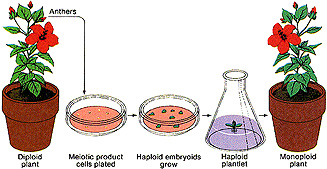
Tehnica de obtinere in culturi de antere sau ovule nu este aplicabila la toate organismele, deoarece exista unele specii refractare la aceste procedee.
O tehnica deosebit de avantajoasa este metoda bulbosum, in care forma diploida de orz (Hordeum sativum) a fost polenizata cu polen de la forma salbatica Hoedeum bulbosum. In succesiunea diviziunilor mitotice cromozomii speciei Hordeum bulbosum au fost eliminati, dand nastere unui embrion haploid. Procesul de haploidizare este aproape similar cu fenomenul de incompatibilitate genetica intre seturile cromozomale ale celor doua specii, chiar daca fecundarea are loc. Prin utilizarea metodei bulbosum s-au obtinut rapid forme homozigote, fiind posibila introducerea in cultura a numeroase varietati noi de orz. Metoda se preteaza si la alte specii.
1. 1. 8. Importanta haploidiei
In studiile de citogenetica, haploizii asigura studierea fiecarui cromozom in parte, evidentiindu-se astfel unele duplicatii, precum si comportamentul cromozomilor in meioza, se pot studia relatiile sinaptice dintre ei. Pe baza imperecherii, sub forma de bivalenti sau multivalenti, s-a stabilit originea poliploida a unor specii.
Importanta genetica a monoploizilor consta in manifestarea integrala a tuturor genelor, inclusiv a celor recesive. Din acest punct de vedere, monoploizii constituie un material ideal pentru studiile de mutageneza. La monoploizi, mutatiile aparute pot fi decelate chiar in anul producerii lor, deoarece se manifesta in fenotip toate genele, inclusiv cele recesive, contrar celor de la poliploizi, care trebuie cautate in generatiile segregante.
Prin dublarea numarului de cromozomi se obtin linii izogene, superioare celor cosangvine.
1. 1. Aloploidia
La formele aloploide are loc multiplicarea seturilor de cromozomi provenite prin hibridare interspecifica sau intergenerica. Formele aloploide se pot realiza prin doua sisteme:
prin incrucisarea dintre specii diploide cu specii poliploide, care au cel putin cate un genom omolog, formand hibrizi indepartati cu grad foarte diferit de fertilitate sau;
prin incrucisarea intre specii indepartate genetic, urmata de dublarea numarului de cromozomi la hibrizii F1, rezulta amfidiploizi sau amfiploizi.
Dupa gradul de omologie a seturilor cromozomale parentale, amfiploizii pot fi segmentali si genomali.
Amfiploizii genomali sunt rezultatul hibridari intre specii cu genomuri complet diferite.
Daca se incruciseaza doua specii diferite A si B rezulta un hibrid AB steril. Prin dublarea numarului de cromozomi, va rezulta un amfiploid AABB, fertil. Amfiploizii genomali prezinta, de regula, fertilitate normala, deoarece imperecherea cromozomilor in meioza se face pe baza omologiei dintre formele parentale sau inrudite, cu care nu se hibrideaza.
![]() Amfiploizi
segmentali
sunt rezultatul hibridarii intre specii cu genomuri partial sau
complet omoloage, urmate de dublarea numarului de cromozomi (AxA' AA
A'A' ). Amfiploizii segmentali au un grad ridicat de
sterilitate, deoarece formeaza figuri bivalente dezordonate (trivalenti
si univalenti), ceea ce duce la separarea haotica in anafaza I a meiozei.
Amfiploizi
segmentali
sunt rezultatul hibridarii intre specii cu genomuri partial sau
complet omoloage, urmate de dublarea numarului de cromozomi (AxA' AA
A'A' ). Amfiploizii segmentali au un grad ridicat de
sterilitate, deoarece formeaza figuri bivalente dezordonate (trivalenti
si univalenti), ceea ce duce la separarea haotica in anafaza I a meiozei.
Autoploizii rezulta atunci cand amfidiploizii isi dubleaza numarul de cromozomi:
![]() (AABB AAAA BBBB).
(AABB AAAA BBBB).
1. 1. 10. Importanta poliploidiei
Pentru natura, poliploidia, in special cea genomala, este deosebit de importanta, asigurand evolutia speciilor.
Un poliploid natural, este graul comun (Triticum aestivum 2n = 6x = 42). Prin studiul speciilor salbatice diploide sau tetraploide, cu numarul de baza, x = 7, s-a reconstituit evolutia graului. In meioza, in celulele mama se formeaza 21 de bivalenti, fiecare cromozom fiind reprezentat de un singur omolog. Formarea bivalentilor este controlata de gena Ph, situata pe bratul lung al cromozomului 5B. Cu toate ca specia este hexaploida, comportamentul "diploid", simulat de formarea bivalentilor, este asigurat de gena Ph.
Graul comun este originar in trei specii diploide diferite si cu genomuri diferite, AA, BB si DD (vezi figura 2.).
Fig. 2.
Diagrama evolutiei posibile a graului hexaploid (Triticum aestivum)
Triticum monococcum 2n=14 genom AA Aegilops mutica 2n=14 genom BB
X
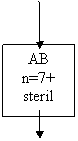
AMFIDIPLOIDIZARE
![]()
Agilops squarosa 2n=14 Genom DD Triticum turgidum 2n=28 Genom AABB
X
![]()
ABD n=7+7+7 steril
![]()
AMFIDIPLOIDIZARE
![]()
Triticum aestivum 2n=42 Genom AABBDD
Originea aloploida a fost stabilita pentru multe alte specii. Prunus domestica este un aloploid intre Prunus spinosa (2n=4x=32) si Prunus cerosifera (2n=2x=16). Alte exemple de specii aloploide: Nicotiana tabacum, 2n = 48 (n. silvestrys, 2n = 24 x N. tomentosiformis, 2n=24), Brassica napus, 2n = 38 (B. oleracea, 2n = 18 x B.capestris, 2n = 20) etc.
Amfiploizii manifesta atat caracteristicile celor doua forme parentale, cat si a poliploizilor, manifestand homeostazie ridicata.
Amfiploidia este o cale deosebit de facila pentru obtinerea de specii noi, artificiale. O prima incercare in acest sens este a lui G.D. Karpecenko (1928), care a obtinut un hibrid fertil intre varza (Brassica oleracea, 2n = 18) si ridiche (Raphanus sativus, 2n = 18). Hibridul obtinut a fost insa steril.
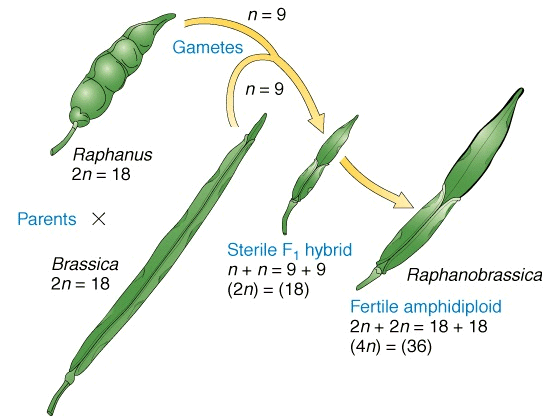
Obtinerea amfiploizilor prin hibridare somatica
Eforturile cercetatorilor in domeniul ameliorarii plantelor sunt focalizate in obtinerea hibrizilor interspecifici pe cale somatica, aceasta pentru a evita dificultatile legate de metodologia clasica (specii refractare, sterilitate etc.).
Metoda hibridarii somatice asigura formarea unor hibrizi a caror forme parentale sunt specii indepartate filogenetic. Se utilizeza in hibridare suspensii de protoplasti de la cele doua forme parentale (celule lipsite de perete celular prin digestie enzimatica). Cele doua suspensii de protoplasti se amesteca in prezenta polietilenglicolului, care stimuleaza fuziunea. Se obtin astfel hibrizi somatici care inglobeaza intr-o celula, atat materialul nuclear, cat si cel citoplasmatic al ambelor forme parentale.
Singurul amfiploid de interes economic este Triticale, un amfiploid obtinut prin hibridare intre grau (Triticum aestivum, 2n=6x=42 sau T. durum, 2n=4x=28) si secara (Secale cereale, 2n=2x=14)(I. Gaspar si Gallia Butnaru, 1984).
Triticale manifesta excelent recombinarea caracteristicilor parentale, randamentul graului cu rusticitatea secarei.
1. 2. ANEUPLOIDIA
Aneuploidia, este fenomenul prin care modificarea numerica afecteaza numai anumite perechi omoloage de cromozomi, adica in nucleul celulelor exista in plus sau in minus, unul sau mai multi cromozomi.
Daca intr-un cariotip alaturi de numarul diploid de cromozomi (2n), se afla unu sau mai multi cromozomi, aneuploidia este de tip hiperploid (polisomie). Daca din garnitura diploida se pierde unul sau se pierd mai multi cromozomi, aneuploidia este de tip hipoploid (oligosomie).
Cu toate ca aspectele genetice si citogenetice ale fenomenului de aneuploidie sunt diferite, in esenta toate au cauze comune si anume:
monodisjunctia cromozomilor in mitoza sau meioza si, in consecinta, repartizarea inegala a materialului genetic la celulele fiice;
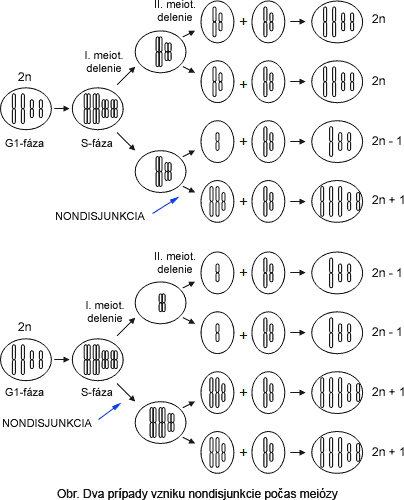
- formarea trivalentilor si univalentilor in loc de bivalenti normali;
- aparitia aberatiilor in structura unor cromozomi;
- absenta sinapsei sau sinapsis partial intre unii cromozomi, fenomen mai frecvent observat la cromozomii lungi.
1. 2. 1. Tipuri de aneuploidie
Hiperploidia sau polisomia, se intalneste in situatia cand, pe langa perechea de cromozomi omologi se gaseste un cromozom in plus, adica 2n + 1, fenomen numit trisomie sau, cand unei perechi de cromozomi i se alatura doi cromozomi, 2n + 2, fenomen denumit tetrasomie.
Trisomia a fost evidentiata, pentru prima data, de A.F. Blakeslee (1921) intr-o cultura de Datura stramonium, 2n = 24, a caror plante posedau habitus modificat, viguroase si cu frunze deosebite de celelalte. J. Belling (1924), analizandu-le citologic constata existenta unui cromozom in plus, adica 2n=24+1, fata de forma euploida normala.
Dupa tipul extracromozomului si a configuratiilor din profaza I, exista mai multe clase de trisomie:
- trisomia primara, atunci cand in meioza extracromozomul se comporta asemanator cu cei doi omologi la care se ataseaza, formand o configuratie in lant, un trivalent. Daca se formeaza un extravalent, la doua perechi de cromozomi, fenomenul se numeste trisomie primara dubla (2n+1+1);
- trisomia secundara, extracromozomul este un izocromozom (izocromozomul se formeaza ca urmare a diviziunii transversale si nu longitudinale a centromerului),care in meioza, in zigonem, formeaza configuratii de inel;
- in trisomia tertiara, extracromozomul este un cromozom translocat, care in meioza impreuna cu celelalte doua perechi, formeaza un lant.
Tetrasomia a fost observata pentru prima oara tot la Datura stramonium. In urma fecundarii unui ovul n+1 cu polen n+1, rezulta un tetrasom. Polisomia poate afecta oricare pereche de cromozomi, atat la organismele diploide cat si la cele poliploide. In general, tetrasomia nu are afecte negative asupra organismului vegetal.
Si la om sunt cunoscute cateva maladii genetice determinate de fenomenul de trisomie autozomala sau heterozomala.
Sindromul Down sau trisomia 21, a fost primul exemplu de trisomie la om, descrisa in 1866 de catre Down. Persoanele afectate au expresia fetei de tip mongoloid, insotita de anomalii cardiace, intarziere minatala, talie redusa, greutate corporala mica.
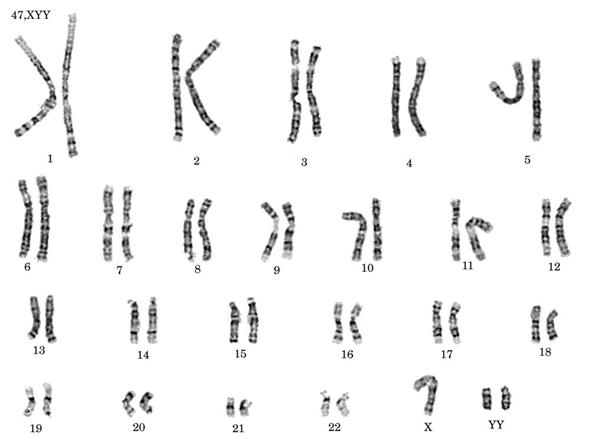
Sindromul Klinefelter (XXY) sau trisomia pentru cromozomii sexului, determina la persoanele afectate o slaba dezvoltare sexuala, atrofie testiculara, ginecomastie, retardare mintala si sterilitate.
Exista si tipuri de sindrom Klinefelter cu constitutia XXYY sau XXXY. Aditia suplimentara de heterozomi amplifica severitatea simptomelor, manifestari antisociale si comportament agresiv, anomalii ale personalitatii, inaltime foarte mare.
Hipoploidia sau oligosomia reprezinta starea unor celule sau a unor organisme care au pierdut un cromozom din garnitura normala, fenomen denumit monosomie (2n-1) sau o pereche de cromozomi, fenomen denumit nulisomie (2n-2).
Hipoploidia se intalneste foarte rar la speciile diploide. Organismele monosomice sau nulosomice nu sunt viabile daca cromozomul care lipseste este de importanta vitala. Sansa de supravietuire o au numai speciile poliploide, ca urmare a fenomenului de compensare, determinat de efectul de doza. Monosomia pentru cromozomul X, sau sindromul Turner, se intalneste la specia umana. Femeile cu un singur cromozom X nu poseda cromatina sexuala, au statura mica, gat plat, dezvoltare sexuala rudimentara, intarziere mintala.
La mamifere, formele trisomice sau monosomice pentru heterozomii X, pot fi depistate citologic prin intermediul cromatinei sexuale Barr.
1. 2. 2. Caracteristicile aneuploizilor
In general, vitalitatea si fertilitatea aneuploizilor, este mai redusa.
Gametul femel tolereaza mai bine aneuploidia, gametii masculi aneuploizi nu sunt viabili, aneuploidia transmitandu-se pe linie materna.
Ca urmare a efectului de doza, polisomii prezinta unele particularitati, specifice cromozomilor suplimentari. La oligosomi, ca urmare a reducerii dozei de material genetic, apar caracteristici noi fata de formele initiale 2n, fapt care permite stabilirea importantei fiecarui cromozom.
1. 2. 3. Importanta aneuploizilor
Din punct de vedere teoretic, aneuploizii asigura identificarea rolului genetic al unor cromozomi, legaturile dintre cromozomi, precum si rolul diferitelor gene in expresia fenotipica a unor caracteristici. Stabilirea rolului pe care-l au cromozomii, prin absenta lor din garnitura cromozomala, este dificil de realizat, deoarece fiecare cromozom are o semnificatie bine conturata in supravietuirea gametului sau a individului.
Desi organismele aneuploide nu au importanta economica directa, cercetarile in acest domeniu au luat amploare, datorita rolului lor in procesul de ameliorare a plantelor.
O realizare deosebita in acest domeniu a avut-o E.R. Sears (1953), care a reusit sa creeze 21 de linii nulisomice la soiul de grau Chinese Spring, stabilind astfel rolul genetic al fiecarui cromozom. Un caracter, daca este controlat de o gena plasata pe un anumit cromozom, efectele acestei gene pot lipsi la plantele nulisomice pentru perechea respectiva de cromozomi.
Dupa obtinerea tuturor liniilor nulisomice si precizarea rolului genetic al cromozomilor s-a putut trece la substituirea acestora dintr-un genom, cu cromozomi apartinand altui genom, de la alta specie. Metoda permite inlocuirea unuia sau mai multor cromozomi din genomul diploid a unei specii, cu un numar egal de cromozomi omologi, proveniti de la o alta specie, realizandu-se astfel genotipuri cu o valoare de recombinare imbunatatita.
In liniile de substiututie intraspecifica, pot fi fixate anumite caracteristici valoroase de la donori cu gene particulare valoroase in receptori cu structura genetica deficitara pentru o gena (rezistenta la boli si la conditii nefavorabile de mediu).
Lucrarile lui B.C. Jenkins (1956), privind aditia si substitutia de cromozomi de la secara (specie donor), purtatoarea unor caractere valoroase (rezistenta la ger, la soluri acide si daunatori) la grau (specie receptor), a dus la obtinerea de linii de substitutie cu 2n=42 cromozomi, dintre care 40 de cromozomi ai graului si doi cromozomi de la secara, purtatori de gene valoroase.
Linii de aditie la grau au fost obtinute de J.G. O'Mara (1941), prin incrucisarea amfidiploidului Triticale (2n=56) cu Triticum aestivum (2n=42). In descendenta au fost identificati indivizi cu 2n = 42 + 2, din care 42 de cromozomi proveneau de la grau si doi cromozomi de la secara.
Utilizarea aneuploidiei in ameliorarea plantelor are si unele dezavantaje, deoarece, uneori specia receptoare pierde gene valoroase, situate pe cromozomul substituit, iar o data cu incorporarea cromozomului strain, pe langa gene valoroase, pot fi incluse si unele gene nedorite.
Numarul de cromozomi, este o caracteristica de baza a tututror organismelor.
Intregul genom (euploidie), sau numai unele perechi de cromozomi (aneuploidie), pot fi modificate de variatiunile numerice ale cromozomilor.
Multiplicarea numarului de cromozomi duce la formarea de genotipuri poliploide. Autoploidia, se manifesta in cazul cand, numarul propriu de cromozomi este multiplicat. La plante, frecvent este intalnita autoploidia si aproape este inxistenta la animale. Poate aparea in mod natural sau poate fi indusa in urma tratamentului cu colchicina sau cu alti agenti care influenteaza diviziunea celulara.
Aloploidia apare prin incrucisarea a doua specii diferite, urmata de dublarea numarului de cromozomi. Aceasta tehnica isi gaseste aplicabilitatea in ameliorarea plantelor cultivate, deoarece permite crearea unor specii noi, care sa imbine proprietatile ambelor forme parentale.
Plantele poliploide prezinta unele caracteristici utile din punct de vedere economic, mai ales atunci cand prezinta interes masa vegetativa.
Aneuploizii au avut de asemenea un rol important in evolutia speciilor. Monosomii (2n-1) si trisomii (2n+1) permit precizarea rolului unor cromozomi si intocmirea hartilor cromozomale.
| Contact |- ia legatura cu noi -| | |
| Adauga document |- pune-ti documente online -| | |
| Termeni & conditii de utilizare |- politica de cookies si de confidentialitate -| | |
| Copyright © |- 2025 - Toate drepturile rezervate -| |
|
|
|||
|
|||
|
|||
Analize pe aceeasi tema | |||
|
| |||
|
|||
|
|
|||