|






 |  |

| Biologie | Botanica | Chimie | Didactica | Fizica | Geografie |
| Gradinita | Literatura | Matematica |
Biologie
|
|
Qdidactic » didactica & scoala » biologie Diviziunea celulara |
Diviziunea celulara
Diviziunea celulara
Diviziunea celulara este cea mai spectaculoasa si mai complexa faza a ciclului celular, in care are loc reorganizarea tuturor componentelor celulare si care cuprinde:
- mitoza sau diviziunea nucleara - in care are loc condensarea, segregarea si distributia materialului genetic celor doua celule fiice;
- citokineza sau diviziunea citoplasmei - care incheie mitoza si permite separarea celor doua celule fiice.
In functie de modificarile care au loc in timpul acestei faze se cunosc doua tipuri de diviziune celulara:
- diviziunea directa sau amitoza, in care nu se formeaza fus de diviziune;
- diviziunea indirecta, in care se formeaza fusul de diviziune, prin intermediul caruia cromozomii vor fi distribuiti in cele 2 celule fiice.
La randul ei, diviziunea indirecta se clasifica in functie de numarul de cromozomi distribuit celor doua celule fiice in doua tipuri:
- tipica sau ecuationala (mitoza) - caracteristica celulelor somatice.
- alotipica sau reductionala (meioza) - caracteristica celulelor sexule, respectiv gametilor.
Ambele tipuri de diviziuni sunt obligatoriu precedate de dublarea cantitatii de ADN.
Indiferent de tipul de diviziune, in acest proces se disting doua etape importante:
- stadiul preparativ, in care are loc dublarea cantitatii de AND si sinteza proteinelor bazice, respectiv a nucleoproteinelor;
- stadiul distributiv, in care are loc distribuirea in mod egal a materialului genetic intre celulele fiice.
1. diviziunea directa sau amitoza
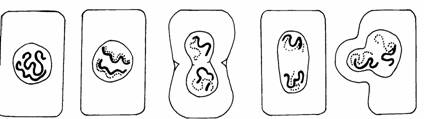
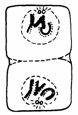
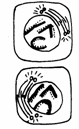
Amitoza (din gr. "a" - fara, "mitos" - fir, filament) reprezinta diviziunea directa a celulei al carei nucleu se afla in interfaza, fiind considerata o forma inferioara de reproducere celulara (Fig. 47).
Inmugurire Clivaj Dublarea cantitatii de ADN
![]()
![]()
Fig. 47. Tipuri de diviziune amitotica (Mariana Sincai, 2000)
Acest tip de diviziune se intalneste la organismele unicelulare atat procariote cat si eucariote: protozoare, metazoare, plante unicelulare si animale nevertebrate.
Amitoza se caracterizeaza printr-o serie de particularitati:
are loc dublarea cantitatii de ADN, urmata sau nu de condensarea cromozomilor;
nu are loc formarea fusului de diviziune;
initial se multiplica nucleolii;
diviziunea nucleului si a citoplasmei are loc prin clivare, strangulare sau inmugurire;
diviziunea nucleului nu este obligatoriu urmata si de diviziunea citoplasmei, ca rezultat obtinandu-se celule multinucleate.
Uneori, din cauza lipsei fusului de diviziune, poate avea loc o distributie inegala a cromozomilor, fapt care conduce la aparitia mutatiilor.
Amitoza este mai frecvent intalnita in celulele din tesuturile patologice (tumori) sau in celulele imbatranite, care si-au redus capacitatea de diferentiere.
In conditii normale, prin amitoza se multiplica unele dintre foitele embrionare ale animalelor, placenta, celulele foliculare ale ovarului, celulele glandelor endocrine, hepatocitele, celulele superficiale ale epiteliului de tranzitie etc.
2. diviziunea indirecta
Diviziunea indirecta este un proces mult mai complex, prin care se realizeaza distribuirea egala a materialului genetic (ereditar) la cele doua celule fiice.
Mitoza sau diviziunea ecuationala
Mitoza (din gr. "mitos" - filament) reprezinta diviziunea indirecta a celulelor somatice, prin care se materialul genetic se distribuie uniform in celulele fiice. Acest tip de diviziune asigura inmultirea celulelor, cresterea si diferentierea individuala, fenomene prin care se asigura continuitatea si variabilitatea genotipului.
Diviziunea mitotica are loc in celulele-mama diploide, cu 2n cromozomi, care in urma procesului de diviziune genereaza doua celule identice cu celula mama si care fiecare va prezenta 2n cromozomi..
Mitoza ocupa aproximativ 10% din ciclul celular si, in functie de specia din care fac parte celulele care se divid, acest proces poate dura intre cateva minute si cateva ore.
In procesul de mitoza intervin o serie de factori, grupati in trei categorii:
factori intracelulari, reprezentati de raportul nucleo-citoplasmatic si nucleolo-nuclear; in consecinta cresterea volumului celular determina diviziunea nucleului si a citoplasmei;
factori intercelulari, reprezentati de fenomenele de interdependenta dintre diversele tipuri de celule (batrane si tinere) din tesut, de numarul optim al populatiei celulare, de relatiile dintre celule si matricea extracelulara, acestia fiind considerati factorii cei mai importanti;
factori generali, reprezentati de factorii externi (temperatura, lumina), aportul unor substantele chimice (vitamine, hormoni) etc.
Mitoza se poate clasifica, dupa gradul de asemanare dintre celula mama si celulele fiice, in urmatoarele tipuri:
mitoza homotipica (mitoza egala)- in care celulele fiice rezultate se aseamana intre ele si cu celula mama, intalnindu-se la celulele nediferentiate (foarte tinere);
mitoza heterotipica - in care celulele fiice se aseamana intre ele, dar sunt mai mature decat celula mama, fiind caracteristica celulelor aflate in procesul de diferentiere;
mitoza homoheterotipica (asimetrica) - in urma careia o celula fiica se aseamana cu celula mama, iar cealalta difera;
mitoza de diferentiere (de intinerire) - in care celulele fiice rezultate sunt mai tinere decat celula mama, fiind specifica limfocitelor.
Perioada dintre doua diviziuni se numeste interfaza. In aceasta faza, nucleul are structura caracteristica, cu cromatina granulara sau dispusa in blocuri mici. La sfarsitul interfazei, prin acumulare de apa, atat nucleul cat si celula se hipertrofiaza si incep procesele de biosinteza ale materialului genetic.
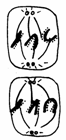
Mitoza cuprinde 4 faze succesive si anume: profaza, metafaza, anafaza si telofaza (Fig. 48).
Telofaza Anafaza Metafaza Profaza Interfaza
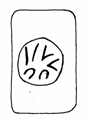
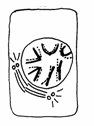


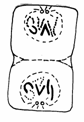
Fig.48. Fazele diviziunii mitotice (Mariana Sincai, 2000)
1. Profaza - este perioada in care cromozomii devin vizibili ca rezultat al spiralizarii, plicaturarii si condensarii cromatinei. Fiecare cromozom se dubleaza longitudinal, evidentiindu-se cele doua cromatide (2 macromolecule de AND), care se dispun rasucit una in jurul celeilalte si se unesc in regiunea centromerului. Spre sfarsitul profazei, nucleolii devin mai mici sau dispar complet, iar membrana nucleara se dezintegreaza. Profaza ocupa aproximativ 50% (30 minute) din mitoza.
Practic, profaza se caracterizeaza prin urmatoarele procese:
dublarea centrului celular, respectiv a centriolilor, inca din interfaza si migrarea acestora spre polii celulei;
formarea fusului de diviziune, in urma polimerizarii microtubulilor din structura centriolilor;
condensarea cromatinei si evidentierea cromozomilor;
dezintegrarea pana la disparitie a nucleolilor, ca urmare a integrarii genelor redundante in cromozomi; partea amorfa a nucleolilor se desprinde si se amesteca cu carioplasma;
dezintegrarea membranei nucleare si a partii nucleare a reticulului endoplasmatic rugos;
amestecul citoplasmei cu sucul nuclear.
Prometafaza este o faza intermediara, care poate fi intalnita suficient de frecvent, si care se caracterizeaza prin:
disparitia membranei nucleare.
initierea interactiunii cromozomilor cu fusul de diviziune. Datorita sistemului motil microtubuli + dineina, cromozomii prezinta miscari oscilatorii, cu ajutorul carora se realizeaza deplasarea acestora spre mijlocul fusului de diviziune.
2. Metafaza - reprezinta faza in care cromozomii sunt bine delimitati si se aranjeaza in regiunea ecuatoriala a celulei, unde formeaza placa metafazica (placa ecuatoriala).
Metafaza este stadiul in care se pot stabili, cu precizie, numarul, forma si marimea cromozomilor specifici fiecarei specii; cuprinde aproximativ 13% (8 minute) din mitoza.
Cromozomii se ataseaza, prin centromer, de filamentele fusului de diviziune (pe fiecare filament cate un cromozom, dispus cu axul longitudinal perpendicular pe acesta).
Cand metafaza se apropie de sfarsit are loc clivarea longitudinala a cromozomilor, respectiv centromerii se scindeaza, de fiecare jumatate ramanand prinsa cate o cromatida, rezultand cromozomii monocromatidici.
In metafaza poate sa continue degradarea membranei nucleare.
|
Asadar, particularitatile metafazei sunt urmatoarele:
aranjarea cromozomilor la ecuatorul celulei;
formarea placii metafazice;
fixarea cromozomilor prin intermediul fusului de diviziune.
3. Anafaza este faza in care are loc migrarea cromozomilor spre polii opusi ai celulei. Astfel, la polii celulei se vor forma doua seturi de cromozomi cu aceeasi constitutie genetica ca si nucleul celulei-mama.
Anafaza reprezinta aproximativ 7% (4 minute) din timpul mitozei si consta in:
deplasarea cromatidelor fiecarui cromozom spre polii celulei (viteza deplasarii este de 0,2-5,0 µm/minut);
aranjarea cromatidelor in timpul deplasarii, conform pozitiei centromerului.
4. Telofaza se caracterizeaza prin formarea la cei doi poli ai celulei a cate un nucleu separat care, dupa despiralizarea cromozomilor devine optic omogen. Spre sfarsitul telofazei apar nucleolii, a caror structura este identica cu cea a celulei mama.
In concluzie, telofaza cuprinde urmatoarele procese:
decondensarea cromozomilor;
formarea unei noi membrane nucleare;
formarea nucleolilor;
disparitia fusului de diviziune;
formarea centrului celular;
citokineza si formarea a doua celule fiice identice cu celula mama.
Citokineza (din gr. "kytos" - cavitate, "kinesis" - miscare) reprezinta diviziunea citoplasmei, respectiv a constituientilor citoplasmatici si ai membranei celulare. Organitele celulare, intr-o masura mai mare sau mai mica, se repartizeaza echilibrat intre celulele fiice.
Vacuolizarea citoplasmei incepe de la ecuatorul celulei, fiind determinata de deplasarea cromozomilor si a organitelor celulare, precum si de distrugerea fusului de diviziune si formarea inelului tranzitoriu. Asamblarea inelului tranzitoriu are loc la inceputul anafazei si este alcatuit din manunchiuri de microfilamente de actina. Inelul tranzitoriu prezinta proprietati contractile. Prin urmare, acest inel se contracta dependent de ionii de Ca, prin mecanismul actina + miozina, micsorandu-si diametrul prin depolimerizarea microfilamentelor de actina.
Structura fusului de diviziune si deplasarea cromozomilor
Fusul de diviziune este alcatuit din filamente centriolo-cromozomiale (filamente cinetocorice) si filamente centriolo-centriolare (filamente polare). La randul lor, fiecare dintre aceste filamente ale fusului de diviziune sunt formate dintr-un manunchi de 100 microtubuli, de care se ataseaza proteine asociate microtubulilor (Fig. 49).
Fusul de diviziune prezinta un rol important in timpul anafazei, cand cromatidele surori se separa si migreaza spre cei doi poli ai celulei.
centriol fus de diviziune Fig. 49. Schema electronooptica a organizarii microtubulilor in fusul de diviziune Inca din
interfaza, centriolii si materialul pericentriolar (impreuna
formeaza centrul celular sau centrozomul) se dubleaza, centriolii
ramanand atasati pe o portiune nucleara pana
la inceputul mitozei. Cel de al doilea centru celular se formeaza prin
asamblarea de tubuline noi, existente in citoplasma, si nu prin
replicarea vechilor centrioli. Dupa migrarea celor doua perechi de
centrioli la polii celulei, acestia declanseaza procesul de
polimerizare a tubulinelor din citoplasma si formarea filamentelor
fusului de diviziune intre cei doi poli ai celulei. cromozomi centriol
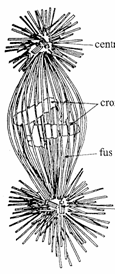
Cinetocorul este o structura de natura proteica, dispusa la nivelul centromerului si care apare sfarsitul profazei. In timpul diviziunii, centromerul fiecarui cromozom prezinta doi cinetocori, dispusi cate unul pe fiecare din cele doua fete opuse ale centromerului (fiecare cromatida din structura cromozomului prezinta cate un cinetocor, ca loc de atasare a filamentelor cinetocorice).
Filamentele cinetocorice sunt formate din manunchiuri de microtubuli care se ataseaza cu un capat de cinetocorul fiecarei cromatide iar celalalt capat radiaza in directia opusa, spre polii fusului de diviziune. Astfel, filamentele cinetocorice prezinta capetele "+" blocate in cinetocori si capetele "-", libere, indreptate spre unul din cei doi centrosomi. Rolul filamentelor cinetocorice este important deoarece servesc la asezarea in pozitie o cromozomilor intre filamentele polare, in placa metafazica si favorizeaza migrarea ulterioara a acestora spre cei doi poli ai celulei.
Filamentele polare din cele doua jumatati ale fusului de diviziune se suprapun partial la ecuatorul celulei.
Deplasarea cromozomilor in anafaza se realizeaza cu ajutorul celor doua tipuri de fibre ale fusului de diviziune.
In faza initiala, numita si anafaza A, deplasarea cromozomilor (dupa clivarea centromerului) se realizeaza cu ajutorul filamentelo cinetocorice, care se scurteaza prin depolimerizarea capetelor libere "-" ale microtubulilor constituienti si care, prin miscarea lor spre poli, atrag cu ele si cromatidele atasate.
In urmatoarea faza, anafaza B, intervin filamentele polare. Astfel, filamentele polare se alungesc, ca urmare a polimerizarii tubulinelor de la capatul lor "+", si aluneca unele peste altele impingand cei doi poli ai celulei, indepartandu-i. Concomitent cu indepartarea polilor celulari, filamentele polare alungite imping si cromatidele separate spre cei doi poli.
Mitoza prezinta o semnificatie importanta din punctul de vedere al biologiei, deoarece prin acest proces se asigura:
constanta numarului de cromozomi;
cresterea si dezvoltarea organismului pluricelular;
integritatea structurala a tesuturilor in caz de pierdere celulara (substituirea eritrocitelor, a celulelor din epiteliul intestinal etc.);
regenerarea tesuturilor si a organelor.
meioza - diviziunea reductionala
Meioza sau diviziunea reductionala (din gr. "meion" - mai putin si "oxis" - conditie) reprezinta diviziunea caracteristica organismelor cu alternanta de faze (haploida - diploida), prin intermediul careia se formeaza, din celule somatice, cei doi gameti (mascul si femel), cu garnitura cromozomiala redusa la jumatate. Cu alte cuvinte, gametii contin un singur cromozom din fiecare pereche, fiind celule haploide (n), care prin contopire, in procesul de fecundare, formeaza celula ou sau zigotul, cu garnitura diploida a speciei.
In meioza, cantitatea de AND se dubleaza o singura data si se imparte de doua ori. Astfel, formarea gametilor este precedata de doua diviziuni succesive deosebite, in care nu se mai produce sinteza ADN:
diviziunea reductionala (primara, heterotipica, meioza I) - prin care se separa cromozomii omologi, fiecare fiind alcatuit din doua molecule de ADN rezultate din replicare. Celulele nou-formate vor primi cate un cromozom intreg din fiecare pereche de cromozomi omologi, prezentand astfel n cromozomi bicromatidici (cu doua molecule de AND identice legate prin centromer);
diviziunea ecuationala (secundara, homotipica, meioza II) reprezinta o mitoza, prin care se separa cromatidele (cele doua molecule de ADN) fiecarui cromozom, rezultand celule cu n cromozomi monocromatidici.
Acest tip de diviziune se intalneste in cadrul ovogenezei si spermatogenezei.
Spre deosebire de mitoza, meioza este un proces mai complex si prezinta o durata mai mare de timp, care poate dura luni sau ani de zile, in functie de specie.
Fiecare din cele doua diviziuni indirecte se desfasoara in patru etape: profaza, metafaza, anafaza si telofaza, cu trasaturile lor caracteristice (Fig. 50).

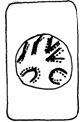
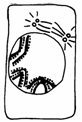

METAFAZA
I PROFAZA
I interfaza I
TELOFAZA TELOFAZA I 2 celule cu 2n cromozomi
![]()

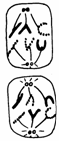

ANAFAZA
I
![]()
Fig.50. Fazele diviziunii meiotice (Mariana Sincai, 2000)
Profaza I este cea mai lunga, ocupand 90% din durata totala de desfasurare al meiozei. Acest lucru se explica prin faptul ca replicarea moleculelor de ADN nu se termina in faza S a ciclului celular, ci se continua si in faza G2 si in profaza.
In profaza I se intalnesc cinci stadii succesive: leptoten, zigoten, pachiten, diploten si diakineza.
In leptoten (din gr. "leptos" - fin, subtire; "teino" - a intinde) are loc condensarea cromatinei si se evidentiaza cromozomii subtiri, formati din doua cromatide, fara clivaj longitudinal. Ei se impletesc intr-o retea numita spirem. De-a lungul cromatidelor se remarca prezenta unor granule intens colorate denumite cromomere, reprezentand zone cu spiralizare mai accentuata (inactiva genetic). In lungul celor doua cromatide, pe una din fete se formeaza o axa proteica lamelara. In leptoten este prezent nucleolul.
In zigoten (din gr. "zigon" - cuplu) are loc conjugarea, respectiv imperecherea cromozomilor omologi (unul matern si unul patern) pe axul longitudinal formand o structura tetracromatidica, numita tetrada (bivalenti). Conjugarea cromozomilor are loc treptat, incepand din anumite puncte situate la extremitati si se extinde asemenea unui fermoar. In acest caz, locii omologi corespund. Daca cei doi cromozomi din tetrada nu sunt identici, conjugarea are loc doar intre segmentele omoloage. Se presupune ca fortele care determina conjugarea pot fi de natura electrostatica sau hidrodinamica. Formarea sinapsei este caracteristica numai meiozei. In zigoten are loc inceputul sinapsei, care se degradeaza in diploten.
Complexul sinaptic sau sinaptonul este o structura proteica axiala, a carei suprafata este inconjurata de fibre radiare, care intra in contact cu membrana nucleara, fiind formata din trei parti:
zona centrala, cu dimensiunea cuprinsa intre 60-120 nm;
un element central, cu dimensiunea cuprinsa intre 10-40 nm, situat de-a lungul celor doi cromozomi omologi conjugati;
doua elemente laterale, cu dimensiuni de 30-40 nm, cate unul pentru fiecare cromozom omolog.
Functia sinaptonului este aceea de a mentine in stransa legatura cromozomii conjugati si de a asigura formarea chiasmelor si a crossing-overului.
In zigoten se sintetizeaza o mica cantitate de ADN, iar prezenta cromomerelor si a nucleolului este o caracteristica morfologica a acestei faze.
In pachiten (din gr. "pachys" - gros) are loc condensarea si spiralizarea cromozomilor conjugati. In aceasta faza, cromozomii sunt mai scurti si mai grosi decat in zigoten si prezinta cromomere. Intre cromatidele nefiice ale cromozomilor omologi se evidentiaza punctele de contact speciale sau chiasmele, considerate expresia citologica a crossing-overului, respectiv schimbul de gene care determina variabilitatea indivizilor din generatiile urmatoare. In pachiten continua sinteza unei cantitati mici de ADN si poate avea loc activarea capacitatii de transcriere a cromozomilor si formarea cromozomilor "perie de lampa". La sfarsitul acestei faze dispar cromomerele.
In diploten (din gr. "diploos" - dublu) incepe clivajul sau separarea cromozomilor pe toata lungimea lor si respingerea cromatidelor cromozomilor omologi, ramanand in contact numai la nivelul chiasmelor. Acest proces incepe in regiunea centromerului. Prezenta chiasmelor reprezinta dovada ca si in aceasta faza are loc crossing-overul. Numarul si distributia chiasmelor sunt caracteristice pentru fiecare bivalent in parte. Scaderea numerica a chiasmelor din fiecare bivalent se realizeaza printr-un proces numit terminalizare, care consta in alunecarea acestora spre extremitatile cromozomilor. Spre sfarsitul diplotenului, sinaptonul se descompune, nucleolii se reduc in dimensiuni, iar la masculi se evidentiaza cromozomii sexuali X si Y (bivalentul sexual X-Y prezinta un aspect liniar).
In diachineza (din gr. "de" - prin, de-a curmezisul; "kinesis" - miscare) cromozomii sunt uniti doar prin cateva chiasme, incepand procesul de detasare de invelisul nuclear. Cele patru cromatide surorii ale bivalentului raman legate prin centromer, iar cele ale omologilor, prin chiasme. Spre sfarsitul acestei faze, nucleolii si membrana nucleara dispar si incepe formarea fusul de diviziune.
In continuare, prima diviziune a meiozei se desfasoara parcurgand aceleasi faze ca si in mitoza:
1. Metafaza I se caracterizeaza prin disparitia membranei nucleare si formarea fusului de diviziune. Cromozomii bivalenti, fixati pe filamentele fusului de diviziune, se indreapta spre ecuatorul celulei formand placa metafazica. In aceasta faza, procesul de terminalizare este incheiat, fiecare bivalent avand cromozomii atasati numai prin chiasmele terminale. Cromozomii din perechi se aseaza intr-o pozitie simetrica: unul este orientat spre un pol, iar altul spre celalalt pol. Filamentele fusului de diviziune se fixeaza de cromozomii din bivalent. La sfarsitul metafazei, intre cele doua centromere ai bivalentului, atasate de filamentele cinetocorice, are loc un fenomen de respingere activa ceea ce conduce la desfacerea din chiasmele terminale a cromozomilor omologi si la indepartarea acestora.
2. Anafaza I se caracterizeaza prin deplasarea spre polii celulei a cate unui cromozom (format din doua cromatide) din fiecare bivalent. Cromatidele cromozomului se deplaseaza perechi, deoarece nu s-a produs diviziunea centromerului. Ca rezultat, la polii celulei are loc separarea setului diploid de cromozomi 2n, in doua seturi haploide n.
3. Telofaza I se caracterizeaza prin formarea a doi nuclei cu seturi haploide (n), respectiv prezinta numarul de cromozomi redus la jumatate. Fiecare cromozom este bicromatidic (contine doua cromatide). Nucleii isi restabilesc structura si se formeaza membrana nucleara. Ulterior, are loc citokineza, prin care se obtin doua celule fiice mai mici.
Dupa telofaza I urmeaza interfaza (interkineza). Aceasta faza nu este intotdeauna obligatorie, la unele organisme putand sa lipseasca chiar si telofaza I si anafaza I.
Dupa interkineza, in care numarul de cromozomi nu se schimba (nu se mai dubleaza cantitatea de ADN) urmeaza a doua diviziune meiotica - diviziunea ecuationala sau homotipica, care se aseamana cu cea mitotica, insa se divid doua celule haploide, care vor formeaza patru celule haploide, cu cromozomi monocromatidici (alcatuiti dintr-o singura cromatida).
Aceasta diviziune se deruleaza, de asemenea, in patru faze: profaza II, metafaza II, anafaza II, telofaza II.
In profaza II are loc condensarea cromozomilor, dezintegrarea nucleolilor si a membranei nucleare si incepe formarea fusul de diviziune.
In metafaza II se termina procesul de formare a fusului de diviziune. Cromozomii se aranjeaza la ecuator, formand placa metafazica, ei fiind fixati in regiunea centromerului de filamentele fusului de diviziune.
In anafaza II cromatidele surori ale fiecarui cromozom se despart si se indreapta spre polii celulei.
In telofaza II, cromozomii, ajunsi la cei doi poli, se despiralizeaza. Pe parcursul acestei faze se reasambleaza nucleii, se formeaza membrana celulara si rezulta patru celule haploide. Aceste patru celule nou formate reprezinta precursorii gametilor.
Importanta biologica a meiozei deriva din urmatoarele consideratii:
meioza asigura constanta numarului de cromozomi in cadrul reproducerii sexuate, micsorand de doua ori numarul acestora in gameti;
In urma fecundatiei, in celula ou sau zigot, se restabileste de fiecare data numarul diploid de cromozomi;
ca rezultat al crossing-overului, prin meioza, se asigura diversitatea sau heterogenitatea genetica. Astfel, populatiile organismelor devin mai heterogene si se pot adapta mai usor la conditiile mediului.
Gametogeneza
Gametogeneza reprezinta procesul prin care are loc formarea celulelor sexuale, respectiv a gametilor celor doua sexe (ovulul - la sexul femel si spermatozoidul - la sexul mascul).
Spermatogeneza
La masculi, spermatogeneza cuprinde mai multe tipuri celulare si diviziuni, grupate in trei etape: multiplicare, crestere si maturare. Acest proces are loc in epiteliul tubilor seminiferi din testicule si incepe odata cu instalarea pubertatii, dupa care se manifesta continuu.
Etapa de multiplicare. Din celulele germinative primordiale deriva spermatogoniile, care reprezinta celulele "cap de serie" ale ciclului spermatocitogenic, prezentand capacitatea de a se divide, prin mitoza, toata viata. La pubertate, in urma unei succesiuni de diviziuni mitotice, spermatogoniile determina aparitia a doua tipuri de celule germinative: spermatogoniile A si B.
Spermatogoniile A sunt celule stem ale liniei seminale, contin un set diploid de cromozomi (2n) dar sunt imature. Dupa cateva diviziuni mitotice o parte din spermatogoniile A se vor diferentia in spermatogonii B care, de asemenea, contin un set diploid de cromozomi.
Spermatogoniile B intra in etapa de crestere, dupa care, prin diviziune mitotica, dau nastere spermatocitelor primare (de ordinul I). Acestea intra in prima etapa a diviziunii meiotice, respectiv in profaza diviziunii reductionale, in care cromozomii omologi sufera fenomene de crossing-over.
Prin diviziunea reductionala a unui spermatocit primar se separa cromozomii omologi si rezulta doua spermatocite secundare (de ordinul II), care contin un set haploid de cromozomi bicromatidici.
Dupa o interfaza foarte scurta, cele doua spermatocite secundare intra in a doua etapa a meiozei, diviziunea ecuationala, prin care se separa cromatidele surori ale fiecarui cromozom. Rezulta astfel patru spermatidii care contin un set haploid, cu n cromozomi monocromatidici.
Ulterior, spermatidiile, care nu se mai divid, intra in etapa de maturare, in care sufera diferentieri morfologice, in urma carora se transforma in spermatozoizi (n cromozomi monocromatidici - Fig. 51).
In spermatogeneza are loc o distributie completa si succesiva a materialului genetic iar cantitatea de citoplasma se reduce si se pierde, astfel ca spermatozoizii rezultati prezinta o cantitate foarte redusa de citoplasma. Prin urmare spermatozoidul este o celula mica, care prezinta un cap ocupat in majoritate de nucleul haploid si o coada reprezentata de flagel - cu rol in motilitate. In capul spermatozoidului, deasupra nucleului se gaseste un rest de vezicula golgiana - acrosomul, care contine enzime litice ale membranei pelucide care inveleste ovulul.
Dintr-o spermatogonie B se obtin la sfarsitul meiozei 16 spermatozoizi.
Spermatocit I (primar) Spermatocit II (secundar)
cu n cromozomi bicromatidici Spermatidii cu n cromozomi
monocromatidici Spermatozoizi
rezultati prin metamorfoza spermatidiilor Mitoza spermatogoniei Spermatogonie Celula germinala primordiala
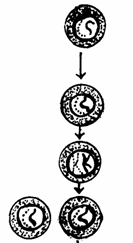
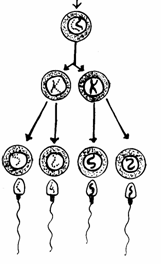
Fig. 51. Stadiile spermatogenezei (Mariana Sincai, 2000)
Pe scurt, procesul de spermatogeneza poate fi redat astfel:
Spermatogonie (2n) → mitoza → spermatocit primar (2n)→ diviziune reductionala → spermatocit secundar (n bicromatidici)→ diviziune ecuationala → spermatidie (n monocromatidici) → transformare → spermatozoid (n).
Ovogeneza
La femele, ovogeneza este initiata in tipul perioadei embrionare, se intrerupe in faza diploten a profazei I si este reluata in viata postnatala, cand apar ciclurile estrale. Procesul de ovogeneza se realizeaza in foliculii ovarieni.
Ovogeneza cuprinde aceleasi etape ca si spermatogeneza, cu cateva particularitati care insa nu schimba sensul procesului general.
Celula stem de la care porneste procesul de ovogeneza este ovogonia, care deriva din celulele germinative primordiale, care in perioada embrionara migreaza in ovarele in curs de formare.
Dupa o perioada de multiplicare prin mitoza, ovogoniile se diferentiaza in ovocite primare sau de ordinul I, cu set diploid de cromozomi (2n), care intra in diviziunea reductionala a meiozei, ramanand in profaza I a acesteia, de la cateva zile la cativa ani, in functie de specie In aceasta perioada de repaus, cromozomii se decondenseaza, are loc o intensificare a activitatii de trascriptionare, astfel ca ovocitele primare isi dobandesc invelisul extern, granulele corticale, isi formeaza ribozomii si ARNm si isi acumuleaza substantele nutritive (proteine, glicogen, lipide) care asigura initierea dezvoltarii embrionare.
La atingerea maturitatii sexuale a specie, sub influenta hormonilor gonadotropi hipofizari, diviziunea reductionala continua si prin separarea cromozomilor omologi, dintr-o ovocita primara rezulta o ovocita secundara - de ordinul II - de talie mare (95% din volumul citoplasmatic total) cu n cromozomi bicromatidici si un globul polar care contine cealalta jumatate de cromozomi bicromatidici si o cantitate redusa de citoplasma (5%).
La majoritatea speciilor, in urma ovulatiei (dehiscienta foliculara) are loc expulzarea ovocitei secundare.
Ovocitele secundare continua meioza intrand in cea de-a doua etapa a acesteia, diviziunea mitotica sau ecuationala, in care are loc separarea in mod egal a cromatidelor surori ale fiecarui cromozom si distribuirea inegala a citoplasmei. Astfel se formeaza ovulul matur, bogat in citoplasma, cu n cromozomi monocromatidici si al doilea globul polar, cu restul de n cromozomi monocromatidici si o cantitate redusa de citoplasma.
Ovulul contine o cantitate mare de vitelus si este invelita la exterior de o membrana pelucida, formata din acid hialuronic, care o protejeaza de anumiti factori mecanici si nu permite patrunderea spermatozoizilor straini de specie sau a altor tipuri de celule (Fig. 52).
Schematic, ovogeneza parcurge urmatoarele etape:
Ovogonie (2n)→ mitoza → ovocite primare (2n)→ diviziune reductionala→ primul globul polar (cu un set de n cromozomi bicromatidici) - structura reziduala + un ovocit secundar (cu n cromozomi bicromatidici) → diviziune ecuationala→ ovul (n monocromatidici) + al doilea globul polar ( cu al doilea set de n cromozomi monocromatidici).
Ovocit I (primar) Ovocit secundar cu n cromozomi bicromatidici Primul globul polar Ovocite primare, care dupa pubertate
continua meioza Al doilea globul polar Diviziune reductionala a
ovocitelor primare Ovul Ovogonie care se divide prin mitoza in
interiorul ovarului Ovogonie diploida Celula germinala primordiala
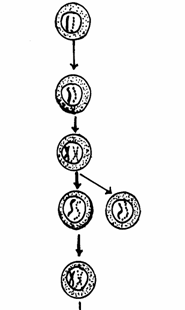
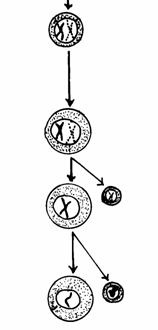
Fig. 52. Stadiile ovogenezei (Mariana Sincai, 2000)
Fecundarea
Fecundarea este procesul esential in fiziologia reproductiei si consta din contopirea celor doua celule sexuale - ovulul si spermatozoidul pentru a da nastere unei celule noi, numita ou sau zigot, la care garnitura de cromozomi va deveni diploida - 2n.
Structura gametilor inaintea fecundarii (fertilizarii)
Fecundarea presupune unele schimbari majore in structura spermatozoidului si ovulului, astfel:
spermatozoidul fecundant trebuie sa recunoasca, sa se ataseze si sa strabata zona pellucida care inconjoara ovulul pentru a se lega de membrana celulara a acestuia;
ovulul trebuie previna in mod activ fecundarea simultana cu mai multi spermatozoizi.
Structura spermatozoidului. Nucleul este continut de capul spermatozoidului care, pentru majoritatea mamiferelor, este turtit sau oval. In timpul spermatogenezei, spermatocitele haploide sintetizeaza coada sau flagelul. De asemenea, mitocondriile se aliniaza helicoidal in jurul portiunii proximale a cozii, formand astfel piesa intermediara. Toate aceste structuri sunt acoperite de membrana celulara. O alta structura a spermatozoidului matur care detine un rol important in procesul de fecundare este reprezentata de acrosom. Aceasta structura reprezinta, in esenta, un lizozom gigant care se formeaza in jurul portiunii anterioare a nucleului. Acrosomul este inconjurat de o membrana care prezinta doua fete: membrana acrosomala interna, orientata spre nucleu si membrana acrosomala externa, care se gaseste in legatura stransa cu plasmalema.
Structura ovulului. La majoritatea mamiferelor, la procesul de fecundare participa ovocitul secundar care, impreuna cu primul globul polar se formeaza in urma primei diviziuni meiotice (diviziunea reductionala). Aceste tipuri celulare: ovocitul secundar si primul globul polar, sunt inconjurate la exterior de un "scut" glicoproteic, numit zona pellucida. Din punct de vedere genetic, ovocitul secundar care ajunge in oviduct se gaseste in metafaza celei de a doua diviziuni meiotice (diviziunea ecuationala). O caracteristica structurala a ovocitului care poate constitui un punct critic in procesul de fecundare este reprezentata de un set de granule corticale. In timpul ovogenezei, ovocitul sintetizeaza mii de granule mici, care se vor acumula in corticala citoplasmei, imediat sub membrana celulara, de care se ataseaza. Granulele corticale ale ovocitelor reprezinta structurile omoloage sau complementare acrosomului din spermatozoizi.
Etapele fecundarii
Fecundarea reprezinta mai mult decat un fenomen izolat, fiind constituita, de fapt, dintr-o inlantuire de evenimente. Intreruperea oricarui eveniment al acestei inlantuiri va produce mai mult ca sigur esecul in realizarea fecundarii. Inceputul desfasurarii acestei inlantuiri de evenimente este reprezentat de o serie de modificari structurale ale spermatozoizilor. Un proces reusit de fecundare presupune nu numai fuzionarea ovocitului cu spermatozoidul dar si prevenirea fuzionarii acestuia cu mai multi spermatozoizi (Fig. 53). Inevitabil, fecundarea ovocitului de catre mai multi spermatozoizi, fenomen denumit polispermie, produce moartea embrionara timpurie.
Fecundarea parcurge urmatoarele etape:
1. Capacitatia spermatozoizilor. In mod normal, spermatozoizii din ejaculatul proaspat sunt incapabili sau foarte putin capabili sa realizeze fecundarea, astfel ca aceste celule trebuie sa parcurga mai intai o serie modificari, denumite generic "fenomen de capacitatie". Acest fenomen este de obicei asociat cu indepartarea proteinelor plasmatice seminale atasate de membrana celulara a spermatozoizilor, reorganizarea lipidelor si proteinelor din structura acesteia, aparitia unui influx de calciu extracelular, o crestere a AMP ciclic si o scadere a pH-ului intracelular. La nivel molecular, capacitatia difera in functie de specie. Capacitatia se declanseaza in timpul transportului spermatozoizilor in tractul genital femel si variaza ca durata in functie de specie (de obicei dureaza cateva ore). Spermatozoizii multor specii de mamifere, inclusiv omul, pot fi capacitati prin incubarea acestora in diferite medii de fertilizare. Prin parcurgerea fenomenului de capacitatie, spermatozoizii se activeaza, manifestand hipermotilitate. Rolul cel mai important al capacitatiei pare a fi acela ca destabilizeaza membrana celulara a spermatozoizilor, pregatindu-i astfel pentru reactia acrozomala.
2. Interactiunea spermatozoid - zona pellucida a ovocitului. Atasarea spermatozoizilor de zona pellucida a ovocitului reprezinta de fapt o interactiune ligand + receptor, cu un grad inalt de specificitate de specie. Fractiunile glucidice ale glicoproteinelor din structura zonei pellucida functioneaza ca receptori pentru spermatozoizi. Moleculele de suprafata ale spermatozoizilor nu sunt cunoscute inca, dar exista anumite proteine care pot indeplini rol de ligand.
3. Reactia acrosomala. Aceasta reactie presupune strabaterea zonei pellucida de catre spermatozoidul care prezinta acrosom (un lizozom modificat, de talie mare, incarcat cu enzime digestive). Aceleasi proteine ale zonei pellucida care initial au servit drept receptori produc o serie de evenimente care conduc la fuzionarea in mai multe puncte a acesteia cu membrana acrosomala externa a spermatozoidului. Aceasta fuzionare si formarea unei vezicule, care reprezinta practic un fenomen de exocitoza, expune continutul acrosomal, conducand la dispersia enzimelor cu rol digestiv. Pe masura ce reactia acrosomala progreseaza si spermatozoidul patrunde in zona pellucida cu atat mai mult se pierde continutul acrosomal, iar membrana acrosomala interna se denudeaza. Spermatozoizii care isi pierd acrosomul inaintea apropierii de ovocit sunt incapabili sa interactioneze cu zona pellucida, fiind, prin urmare, incapabili sa realizeze fecundarea.
4. Strabaterea zonei pellucida. Forta propulsiva constanta exercitata de coada flagelata in combinatie cu enzimele acrosomale permit spermatozoidului sa creeze o "poarta" prin zona pellucida. Prin urmare, acesti doi factori: motilitatea si enzimele digestive, permit spermatozoidului sa traverseze zona pellucida.
5. Interactiunea spermatozoid - ovocit. Dupa ce a strabatut zona pellucida, spermatozoidul se ataseaza si fuzioneaza cu membrana celulara a ovocitului. Atasarea se realizeaza in regiunea posterioara sau post-acrosomala a capului spermatozoidului. Spermatozoizii activati contin in membrana lor proteine acide - fertiline, iar ovulele contin in membrana glicoproteine - fertilizine, reactia de cuplare fiind asemanatoare reactiei antigen + anticorp.
6. Activarea ovocitului si reactia corticala. Inaintea fecundarii, ovocitul se gaseste intr-o stare de quiescenta, "arestat" in metafaza celei de a doua diviziuni meiotice. Imediat inaintea interactiunii cu spermatozoizii, ovocitul sufera numeroase modificari metabolice si fizice care poarta numele generic de "activarea ovocitului". Efectele predominante ale acestei activari includ cresterea concentratiei intracelulare a ionilor de calciu, finalizarea celei de a doua diviziuni meiotice si asa numita reactie corticala, care se refera la o exocitoza masiva a granulelor corticale. Aceste granule contin numeroase enzime, inclusiv anumite proteaze, care, consecutiv exocitozei, difuzeaza in zona pellucida si / sau interactioneaza cu membrana celulara a ovocitului.
7. Reactia zonei pellucida. Aceasta reactie se refera la alterarea structurii zonei pellucida, catalizata de proteazele continute de granulele corticale. La majoritatea mamiferelor, importanta reactiei zonei pellucida este reprezentata de blocarea instalarii polispermiei. Acest efect este rezultatul a doua modificari semnificative induse in zona pellucida:
- consolidarea zonei pellucida, care impreuna cu membrana celulara a ovocitului devin impermeabile pentru alti spermatozoizi;
- distrugerea receptorilor pentru spermatozoizi din structura zonei pellucida.
Realizarea in conditii normale a fecundatiei este conditionata de o serie de factori favorizanti, printre care calitatea ovulului si a spermatozoidului joaca rolul esential.
Capul nuclear al spermatozoidului incorporat devine pronucleu mascul. Apoi are loc diviziunea mitotica a ovocitei secundare, cu eliminarea celui de-al doilea globul polar iar setul cromozomial al ovulului ramas in citoplasma se grupeaza si se delimiteaza printr-o membrana, transformandu-se in pronucleu femel.
Pronucleii sunt antrenati prin contractia microfilamentelor de actina spre centrul ovulului, cresc in volum si fuzioneaza, realizand amfimixia. Ca urmare, seturile haploide de cromozomi ale celor doi pronuclei se vor combina, realizandu-se diploidia noii celule formate.
Celule foliculare
Dupa realizarea fecundarii, pronucleii celor doua
celule sexuale isi inceteaza existenta, luand nastere noua
celula, oul sau zigotul.
Membrana pelucida Amfimixia Spermatozoid Fuziunea membranei spermatozoidului cu membrana
ovulului Penetrarea zonei pelucida Acrosom Ovul Nucleu
![]()
![]()
![]()
![]()
![]()
![]()
![]()
![]()
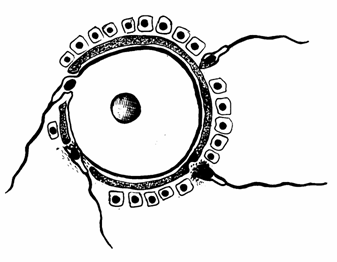
Fig. 53. Actiunea acrosomului in procesul de fecundare al celor doi gameti (Mariana Sincai, 2000)
| Contact |- ia legatura cu noi -| | |
| Adauga document |- pune-ti documente online -| | |
| Termeni & conditii de utilizare |- politica de cookies si de confidentialitate -| | |
| Copyright © |- 2025 - Toate drepturile rezervate -| |
|
|
|||
|
|||
|
|||
Referate pe aceeasi tema | |||
|
| |||
|
|||
|
|
|||