|






 |  |

| Biologie | Botanica | Chimie | Didactica | Fizica | Geografie |
| Gradinita | Literatura | Matematica |
Biologie
|
|
Qdidactic » didactica & scoala » biologie Receptori si mesageri |
Receptori si mesageri
Receptori si mesageri
1. mesagerii sau liganzii
Mesagerii sau liganzii sunt substante chimice care intervin in metabolismul celular, ei actionand asupra unor receptori celulari specifici aflati in plasmalema, in citosol sau in membrana nucleara si care actioneaza in concentratie foarte mica (in jur de 10-8 M).
In raport cu distanta la care actioneaza, pot fi clasificati in trei grupe: endocrini, paracrini si autocrini (Fig. 17).
Mediatori chimici locali Neurotransmitatorii care
actioneaza asupra celulelor adiacente Hormoni ![]()
![]()
![]()
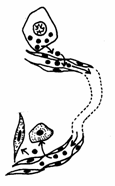
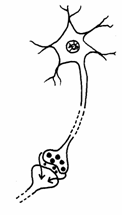
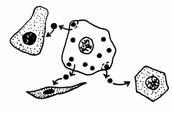
Fig. 17. Tipuri de mesageri: endocrini, paracrini, autocrini (Mariana Sincai, 2000)
Mesagerii endocrini sunt substante eliberate direct in sange de catre celulele glandelor endocrine si care se numesc hormoni. Actiunea hormonilor se realizeaza de regula asupra unui grup de celule tinta aflate la distanta.
Mesagerii paracrini actioneaza numai asupra celulelor adiacente, care sunt in directa legatura cu celulele care i-a elaborat. Astfel, transmiterea impulsului electric de la o celula nervoasa la alta sau la o celula musculara se realizeaza prin intermediul unor semnale chimice numite neurotransmitatori.
Mesagerii autocrini sunt substante elaborate de anumite celule care, actioneaza, 'in revers', chiar asupra celulei care le-a produs sau asupra celulelor din imediata vecinatate. Spre exemplu, multe celule tumorale elaboreaza factori de crestere (TGF - tumor growth factors), care actioneaza atat asupra celulei tumorale insasi cat si asupra celulelor sanatoase vecine, provocand dezvoltarea tumorii.
Celulele endocrine si neuronii sunt celule inalt specializate in semnalizarea chimica, hormonii si neurotransmitatorii fiind mesageri de ordinul I ai acestor celule. Pe cale nervoasa si endocrina se asigura reglarea functiilor tuturor tesuturilor din organism.
Desi, sub unele aspecte, mecanismele semnalizarii chimice si chiar substantele elaborate ca mesageri de celulele endocrine si de neuroni sunt asemanatoare, exista totusi unele deosebiri esentiale. Astfel, hormonii actioneaza la o concentratie foarte scazuta, ei fiind diluati in sange si purtand informatia la o distanta mare, in timp ce neurotransmitatorii actioneaza la concentratii mai mari, transmiterea informatiei fiind mult mai rapida, iar impulsurile electrice sunt transformate in impulsuri chimice la contactul celor doua celule.
Majoritatea liganzilor par sa nu aiba alt rol decat legarea de receptorul specific. Prin urmare, liganzii nu sunt metabolizati, nu prezinta activitate enzimatica si nu intervin direct in activitatea celulara. Singura functie a mesagerilor pare a fi modificarea proprietatilor receptorului; care apoi semnalizeaza celulei ca o anumita substanta este prezenta in mediul din jur.
Celulele tinta pot adesea modifica sau degrada liganzii si astfel pot sa modifice sau sa opreasca raspunsul celulelor la semnalul chimic respectiv.
2. receptorii
Receptorii sunt dispozitive moleculare specializate asezate pe fata externa a membranei celulare, in nucleolema sau in citoplasma, cu ajutorul carora se intercepteaza semnale sau mesaje venite chmice.
Receptorii sunt proteine care au functia de a lega specific si cu mare afinitate anumite substante chimice denumite liganzi sau mesageri. Legatura dintre liganzi si receptorii specifici declanseaza raspunsuri celulare caracteristice. Astfel se produc in organismul viu foarte multe interactiuni intre celulele care secreta liganzi si celulele "tinta".
Dupa originea ligandului pe care il leaga se cunosc doua mari categorii de receptori:
Receptorii pentru substante exogene, din care fac parte receptorii pentru virusuri, toxine bacteriene, antigene straine organismului, anumite medicamente etc.;
Receptorii pentru substante endogene, din care fac parte:
receptorii pentru neurotransmitatori, localizati in membrana postsinaptica a celulelor nervoase, musculare sau a altor celule efectoare. In aceasta categorie se incadreaza receptorii pentru: acetilcolina, dopamina, noradrenalina, serotonina, adrenalina, histamina, glicina, acid gamma-aminobutiric, acid glutamic, acid aspartic care modifica permeabilitatea pentru ioni a membranelor asigurand functionarea sinapselor.
receptorii pentru hormoni, care sunt localizati diferit in celula, in functie de structura chimica a acestora. Astfel, daca hormonul este hidrofob (hormonii steroizi si hormonii tiroidieni) acesta patrunde usor prin plasmalema celulei tinta si se leaga de receptorii intracelulari. In schimb, hormonii hidrofili se leaga de receptorii specifici din plasmalema, care transmit apoi celulei informatia necesara modificarilor metabolice (hormonii cu structura glicoproteica ca insulina, glucagonul, parathormonul, hormonii hipofizari).
receptorii implicati in reactiile imunitare cuprind receptori pentru antigene endogene, receptori pentru anticorpi si receptori pentru complement.
3. EFECTELE CELULARE ALE LEGARII LIGANzilor de receptori
O serie de observatii au relevat faptul ca acelasi receptor poate fi prezent in tipuri foarte variate de celule si ca legand acelasi tip de ligand poate declansa raspunsuri diferite in fiecare tip de celula.
De asemenea, celule foarte diferite raspund diferit la acelasi ligand. Un exemplu in acest sens il constituie receptorii pentru acetilcolina, care se gasesc atat in plasmalema celulelor musculare cardiace si striate, cat si in plasmalema celulelor acinoase pancreatice. Astfel, eliberarea acetilcolinei de catre un neuron adiacent determina contractia celulelor musculare striate, slabirea contractiei miocardocitelor si exocitoza granulelor de secretie din celulele acinoase pancreatice.
|
Pe de alta parte s-a remarcat ca in unele celule, mesageri diferiti declanseaza acelasi raspuns celular. De exemplu, in celulele hepatice atat legarea glucagonului cat si a epinefrinei de receptorii lor specifici induce degradarea glicogenului si eliberarea glucozei in sange.
Legarea liganzilor de receptori se realizeaza prin legaturi de hidrogen, legaturi hidrofobe, intr-o anumita zona specifica din molecula receptorului, complexul ligand+receptor declansand o serie de modificari ale plasmalemei cu efecte intracelulare.
1. Redistribuirea receptorilor si agregarea lor
Legarea liganzilor de receptori determina redistribuirea si agregarea acestora in anumite zone ale plasmalemei, urmata de captarea complexului ligand+receptor in interiorul celulei, prin endocitoza. Ca si exemple in acest sens sunt luate in considerare fagocitoza exercitata de macrofage si pinocitoza colesterolului de catre celulele animale (Fig. 18).
![]()
Spatiu
intracelular Receptori ligand + receptor = agregare Lipide
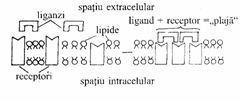
Fig. 18. Agregarea receptorilor din membrana celulara
2. Modificari ale permeabilitatii plasmalemei
Este semnalata in legarea neurotransmitatorilor de receptorii membranei postsinaptice. Se deschid astfel o serie de canale pentru ioni, canale cu poarta comandata de ligand, care inverseaza potentialul de membrana, realizand transmiterea impulsului nervos (potentialul de actiune) neuronului adiacent.
In acest tip de fenomene intervin si mediatori chimici locali care actioneaza asupra celulelor din imediata vecinatate. Ei sunt in general preluati rapid de celule si distrusi la fel de rapid, astfel incat in sange patrund in cantitate relativ redusa. Unii mediatori locali sunt elaborati de celule specializate in aceasta functie. De exemplu, histamina, secretata de mastocite, este eliberata rapid prin exocitoza in cazuri de leziuni, infectii locale sau reactii imunologice. Histamina provoaca local permeabilizarea capilarelor, vasodilatatie si trecerea din sange in tesuturile din jur a anumitor proteine serice, a macrofagelor si a leucocitelor cu rol in raspunsul imun.
Un alt grup de mediatori chimici locali sunt prostaglandinele. Acestea sunt elaborate continuu, nu sunt stocate, au efecte foarte variate si sunt distruse rapid. Prostaglandinele, grupate in noua tipuri chimic distincte, moduleaza raspunsul altor hormoni, stimuleaza contractia musculaturii netede, agregarea trombocitelor si favorizeaza procesele inflamatorii, toate aceste efecte fiind legate si de actiunea lor de permeabilizare a membranei celulare.
3. Activarea unor enzime din plasmalema
Activarea unor enzime din plasmalema este un proces intalnit mai ales in cazul hormonilor hidrosolubili (proteici). Acesti hormoni, desi prezinta efecte rapide, imediate si de scurta durata (cateva secunde) nu pot strabate membrana celulara, ei interactionand cu receptorii de suprafata ai plasmalemei.
Hormonii liposolubili (hormonii steroizi, tiroidieni, acidul retinoic) sunt structurati pe baza de colesterol si ca urmare ei pot strabate membrana celulara si sa se fixeze pe receptori din nucleu sau citosol, prezentand efecte de lunga durata (ore sau zile).
Complexul hormon hidrosolubil si receptor activeaza anumite enzime specifice care declanseaza rapid cresterea concentratiei intracelulare a unui mesager de ordinul 2, respectiv AMPc sau GTPc
Mecanismul activarii enzimelor din plasmalema se bazeaza pe utilizarea de catre receptorii de membrana a unei proteine, care comunica semnalul de la suprafata unei anumite enzime care genereaza un mesager de ordinul II intracelular, respectiv informatia externa este transformata intr-o informatie interna celulei (Fig. 19).
Adenilatciclaza este o proteinenzima de membrana care in portiunea citoplasmatica prezinta structuri de legare a ATP-ului, iar pe fata exoplasmica poseda legaturi receptoare pentru proteine.
A treia proteina de membrana, care functioneaza ca o punte de legatura intre receptor si adenilatciclaza este proteina Gs numita asa datorita legaturilor pe care le poseda pentru GDP si GTP (guanozin difosfat si trifosfat).
Proteina G este alcatuita din trei lanturi peptidice α, β, γ. Subunitatea α, numita si Gsα leaga GTP si GDP si de asemenea se conecteaza la receptorul adenilatciclazei.
GTP legat de proteina Gα este hidrolizat in GDP + p. In momentul legarii GTP de Gsα, aceasta se disociaza de subunitatile Gβ si γ. Subunitatea Gsα cu GTP-ul legat sufera modificari conformationale, devenind capabila sa lege si sa activeze adenilatciclaza. Daca Gsα leaga GDP in loc de GTP nu mai are loc disocierea subunitatilor Gsβ,γ si nu se mai activeaza adenilatciclaza. In celulele nestimulate hormonal, majoritatea moleculelor de Gs sunt legate cu GDP. In momentul cand hormonii actioneaza asupra celulelor si se leaga de receptorii β-adrenergici, le modifica conformatia provocand inlocuirea GDP-ului legat de Gs cu GTP. Astfel are loc o stimulare foarte rapida, dar nu si scurta, a adenilatciclazei deoarece GTP este hidrolizat in cateva secunde in GDP care asociaza din nou Gsα cu Gsβγ determinand inactivarea adenilatciclazei.
Actiunea hormonilor asupra celulelor poate fi in sensul cresterii concentratiei de AMPc sau a scaderii acestuia. De exemplu, in celulele adipoase, epinefrina, glucagonul si ACTH-ul stimuleaza adenilatciclaza, in timp ce prostaglandina PGE1 o inhiba.
In cazul de actiunii de inhibitie a adenilatciclazei intervine proteina G1 (inhibitoare) care este formata din aceleasi subunitati β si γ ca si la Gs dar difera subunitatea α. Astfel, G1 leaga GTP si disociaza in G1α se leaga de adenilatciclaza de care o inhiba.
Hormon
Hormon
![]()
GDP
![]()

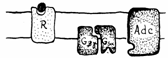
Hormon Hormon
![]()
GTP![]()
![]()
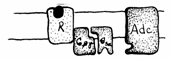

P GTP
![]()
![]()
![]()
![]()
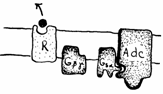
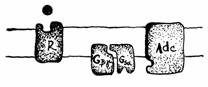
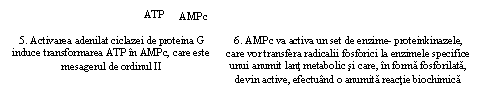
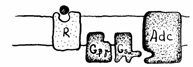
Fig. 19. Efectele legarii liganzilor de receptori si formarea AMPciclic,
mesager de ordinul II (Mariana Sincai, 2000)
O serie de studii au relevat ca unele toxine bacteriene pot modifica ireversibil proteinele G. Astfel este cunoscuta actiunea toxinei holerice, o proteina produsa de bacteria Vibrio cholere, care activeaza ireversibil adenilatciclaza din celulele epiteliale ale intestinului provocand o crestere a nivelului de AMPc si respectiv o diaree masiva, cauzata de trecerea apei din sange in intestinul subtire. Acest fenomen este datorat faptului ca GTP ramane legat de Gsα si nu este hidrolizat la GDP. Astfel, adenilatciclaza este activata continuu, mentinand o concentratie mare de AMPc in celule.
Nivelul de AMPc este controlat prin activarea adenilatciclazei dar si prin mecanismul de degradare al AMPc de catre AMPc - fosfodiesteraza, care sfarseste efectul stimularii hormonale. Multe AMPc - fosfodiesteraze sunt activate ca urmare a cresterii in citoplasma a concentratiei de Ca2+. Ionii de calciu, Ca2+, se leaga se leaga de calmodulina, impreuna AMPc-fosfodiesteraza, pe care o activeaza si, in consecinta, are loc hidroliza AMPc. In unele celule, reglarea nivelului AMPc se realizeaza prin eliberarea lui in mediul extracelular.
4. Mobilizarea ionilor de Ca2+ ca mesageri de ordinul 2
Celulele animale care folosesc ca mesager de ordinul II AMPc sunt foarte numeroase, dar la fel de numeroase sunt si celulele care utilizeaza ionii de calciu.
Nivelul ionilor de calciu in citoplasma este mentinut la o concentratie de 0,2μM, fenomen realizat cu ajutorul pompei Ca2+ATP-aza, care scoate ionii de calciu in mediul extracelular, si ii introduce in canalele reticulului endoplasmatic sau in caveolele citoplasmatice de stocare.
O crestere a concentratiei ionilor de calciu in citoplasma induce o serie de fenomene celulare cum ar fi intensificarea secretiei celulare specifice, degradarea glicogenului, interactiunea actinei cu miozina etc.
Efectele celulare a ionilor de calciu sunt legate de o proteina specifica numita calmodulina, alcatuita din 148 de aminoacizi si care se intalneste in toate celulele eucariote. Ionii de Ca2+ formeaza cu calmodulina un complex care activeaza alte numeroase enzime din diferite procese metabolice. Printre enzimele activate de complexul Ca2+ - calmodulina amintim si AMPc - fosfodiesteraza care degradeaza AMPc. Astfel, cele doua tipuri de mesageri de ordinul II interactioneaza in reglarea metabolismului celular (Fig. 20).
Calmodulina prezinta patru locuri de legare a ionilor de calciu, astfel incat modificari mici ale nivelului calciului citoplasmatic produc mari modificari in concentratia calmodulinei active.
Mesagerii de ordinul II nu numai ca traduc semnalele extracelulare in semnale intracelulare, dar realizeaza si o amplificare foarte mare a semnalului initial. Fiecare ligand care se leaga de un receptor activeaza mai multe molecule de proteina G, deci mai multe molecule de adenilatciclaza. Aceaste din urma enzime, la randul lor, catalizeaza conversia unui mare numar de molecule de ATP in AMPc.
De asemenea, in cazul in care un ligand induce patrunderea Ca in celula, de fapt are loc patrunderea mai multor ioni de acest tip, care se leaga de mai multe molecule de calmodulina. Astfel se activeaza numeroase proteine si enzime, deci au loc adevarate avalanse enzimatice la legarea unui singur ligand.
Ca+2 Activarea enzimei
Calmodulina Ca+2 calmodulina
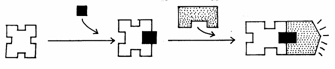
Fig. 20. Ionii de calciu ca mesageri de ordinul II in activarea unor
enzime specifice (Mariana Sincai, 2000)
Datorita acestor mecanisme de amplificare, procentul receptorilor nu depaseste 1 % din totalul proteinelor de membrana, eficienta lor fiind insa foarte mare.
Alti mesageri de ordinul II din citoplasma sunt: l, 4, 5 - inozitoltrifosfatul si l, 2-diacilglicerolul.
O serie de observatii au relevat ca 1, 4, 5- inozitoltrifosfatul actioneaza ca ligand prin deschiderea canalelor cu poarta pentru ionii de calciu din membranele reticulului endoplasmatic, eliberandu-i in citoplasma.
l, 2-diacilglicerolul, ca mesager de ordinul II, determina activarea unei familii de proteinkinaze din plasmalema, cunoscute sub termenul de proteinkinaza C. De fapt, proteinkinaza C este activata atat de 1,2-diacilglicerol cat si de ionii de calciu. In mod normal, proteinkinaza C este o proteina inactiva aflata in citoplasma, care, in prezenta ionilor de calciu, se leaga de plasmalema, unde poate fi apoi activata de 1,2 - diacilglicerol. Proteinkinaza C joaca un ro1 important in procesele de crestere si in metabolismul general al celulelor. De exemplu, in ficat stimularea proteinkinazei C inhiba sinteza de glicogen si faciliteaza degradarea lui in glucoza. O serie de studii privind functiile proteinkinazei C au relevat ca aceasta poate fi activata de unele substante liposolubile numite promotori tumorali, capabile sa transforme o celula normala intr-o celula maligna, cu o crestere necontrolata. Activarea proteinkinazei C este probabil "cheia" evenimentelor proliferative in celula.
Observatii recente au evidentiat ca 1,2-diacilglicerolul si l,4,5- inozitoltrifosfatul se obtin prin hidroliza unui fosfolipid din plasmalema si anume fosfatidil inozitol 4,5 - bifosfat. Hidroliza acestui fosfolipid se realizeaza de catre fosfolipaza C.
1,2-diacilglicerolul ramane in membrana, iar l, 4, 5-inozitoltrifosfatul trece in citoplasma.
Fenomenele de reglare a receptorilor
Cantitatea receptorilor hormonali de la suprafata celulelor nu este constanta. Numarul acestor receptori poate fie sa creasca sau sa scada, permitand celulei sa raspunda optim la schimbari mici ale concentratiei de hormoni.
Numarul receptorilor se poate reduce prin endocitoza sau blocare cu anumiti liganzi. Sinteza de noi receptori este un proces lent care dureaza peste 24 de ore.
| Contact |- ia legatura cu noi -| | |
| Adauga document |- pune-ti documente online -| | |
| Termeni & conditii de utilizare |- politica de cookies si de confidentialitate -| | |
| Copyright © |- 2025 - Toate drepturile rezervate -| |
|
|
|||
|
|||
|
|||
Referate pe aceeasi tema | |||
|
| |||
|
|||
|
|
|||