|






 |  |

| Biologie | Botanica | Chimie | Didactica | Fizica | Geografie |
| Gradinita | Literatura | Matematica |
Chimie
|
|
Qdidactic » didactica & scoala » chimie Sistemul colinergic - acetilcolina |
Sistemul colinergic - acetilcolina
Sistemul colinergic cuprinde acetilcolina impreuna cu totalitatea structurilor care sintetizeaza, utilizeaza si distrug acetilcolina.
Acetilcolina este sintetizata in citoplasma terminatiilor sinaptice ale anumitor neuroni numiti neuroni colinergici, tocmai datorita faptului ca sintetizeaza acetilcolina si o utilizeaza ca neurotransmitator. Sinteza are loc sub influenta enzimei colinacetilaza sau acetilcolintransferaza care favorizeaza formarea unei legaturi esterice intre acidul acetic si colina. Acidul acetic este furnizat in aceasta reactie sub forma de acetil-coenzima A de catre mitocondrii, iar terminatiile presinaptice colinergice sunt bogate in mitocondrii. Colina provine in citoplasma terminatiei colinergice din mediul inconjurator acestei terminatii. Membrana presinaptica este dotata cu doua sisteme de captare a colinei din exteriorul terminatiei presinaptice in citoplasma acesteia, unul rapid si altul lent. Sistemul rapid este influentat de concentratia extracelulara a ionilor de sodiu. Nici unul din cele doua sisteme nu pare sa fie influentat de concentratia intracelulara de acetilcolina. Colinacetilaza sau acetilcolintransferaza este sintetizata la nivelul corpilor celulari ai neuronilor colinergici si migreaza de-a lungul axonului pana la nivelul terminatiilor presinaptice.
colinacetilaza
![]() (CH3)3N+-CH2-CH2-OH + CoA-S-CO-CH3
(CH3)3N+-CH2-CH2-OH + CoA-S-CO-CH3
→ (CH3)3N+-CH2-CH2-O-CO-CH3 + CoA-SH
Fig. nr. 7.1. Sinteza acetilcolinei din colina si radicalul acetil furnizat de acetilcoenzima A, sub influenta
enzimei numita colinacetilaza sau acetilcolintransferaza
Acetilcolina astfel sintetizata este captata in veziculele de depozit presinaptice, printr-un mecanism transportor specific, probabil prin schimb cu ioni de hidrogen. In veziculele de depozit acetilcolina este stocata impreuna cu alti cotransmitatori printre care adenozintrifosfat (ATP), prostaglandine si proteoglican. Se apreciaza ca o vezicula de depozit contine intre 1 000 si 50 000 de molecule de acetilcolina iar intr-o terminatie presinaptica exista probabil peste 300 000 de vezicule de depozit. Masuratorile efectuate pe un singur canal ionic au aratat ca o singura molecula de acetilcolina este capabila sa modifice potentialul membranei celulare cu 3 10-7 V (3 10-4 mV). Aceasta inseamna ca o cuanta de acetilcolina eliberata in fanta sinaptica modifica potentialul membranei postsinaptice cu cel putin 0,3 mV, generand ceea ce se numeste un potential postsinaptic miniatural. In urma depolarizarii presinaptice are loc insa o crestere a influxului de calciu prin membrana presinaptica ceea ce va antrena o succesiune de evenimente care se vor solda cu eliberarea simultana a cca. 100 de cuante de acetilcolina. Acestea vor putea modifica potentialul membranei postsinaptice cu cel putin 30 mV, valoare capabila sa declanseze un potential de actiune propagat.
Acetilcolina eliberata in fanta sinaptica difuzeaza in spatiul sinaptic si se fixeaza de receptorii colinergici postsinaptici exercitandu-si efectul caracteristic. Efectul acetilcolinei este insa repede terminat prin difuziunea substantei in afara spatiului sinaptic si, mai ales, prin metabolizarea sa rapida. Metabolizarea acetilcolinei consta in ruperea legaturii esterice si descompunerea acetilcolinei in cele doua componente, acidul acetic si colina. Acidul acetic este metabolizat pe cand acetilcolina este repede recaptata in citoplasma terminatiei presinaptice pentru a fi reutilizaza la sinteza de noi molecule de acetilcolina. Hidroliza moleculei de acetilcolina poate avea loc sub influenta a doua enzime, una specifica pentru acetilcolina numita acetilcolinesteraza (colinesteraza sau colinesteraza adevarata) si alta mai putin specifica numita pseudocolinesteraza (sau butirilcolinesteraza). Efectul acetilcolinesterazei este nu numai specific dar si foarte rapid, ea putand hidroliza molecula de acetilcolina in mai putin de o milisecunda (cca. 150 microsecunde). Acetilcolinestaraza se gaseste cu precadere in neuronii colinergici, atat in corpul celular cat si in terminatiile dendritice sau axonale, dar si in alte tesuturi. Realizeaza concentratii relativ ridicate in jurul sinapselor colinergice. La nivelul placii motorii acetilcolinesteraza este dispusa foarte strategic pe membrana postsinaptica la periferia sinapsei. Pseudocolinesteraza se gaseste mai putin la nivelul neuronilor cat mai ales la nivelul altor structuri cum ar fi nevrogliile, sangele, ficatul.
Desi metabolizarea acetilcolinei este o cale foarte importanta de inactivare a neurotransmitatorului, nu trebuie neglijat rolul difuziunii acetilcolinei in afara spatiului sinaptic. Acest proces a fost cel care a stat la baza descoperirii acetilcolinei ca neurotransmitator. Initial, in 1921, Otto Loewi a descoperit ca perfuzatul obtinut de la un cord de broasca care a fost stimulat vagal produce la o a doua broasca nestimulata vagal un raspuns identic cu cel obtinut in urma stimularii vagale. Aceasta l-a facut sa constidere ca stimularea vagala elibereaza o anume substanta chimica care a fost numita initial Vagusstoff (substanta-vag) sau parasimpatina. Ulterior, in 1926, Loewi si Navratil au identificat parasimpatina ca fiind acetilcolina. Aceste aspecte istorice demonstreaza ca difuziunea acetilcolinei in afara spatiului sinaptic este atat de importanta incat realizeaza in afara spatiului sinaptic concentratii sufucient de mari pentru a produce efecte caracteristice stimularii sinaptice. Desigur la acumularea si persistenta acetilcolinei in afara sinapselor contribuie de asemenea faptul ca in afara sinapselor nu exista acetilcolinestaraza care sa inactiveze acetilcolina ci cel mult pseudocolinesteraza care are un efect lent.
Efectele acetilcolinei se realizeaza prin intermediul unor receptori specifici numiti receptori colinergici. Efectul acetilcolinei fiind unul specific este evident faptul ca pe receptorul colinergic exista un situs receptor a carui structura sterica trebuie sa fie complementara cu structura sterica a acetilcolinei. Plecand de la structura chimica a acetilcolinei se pot face presupuneri privind structura sterica a situsului receptor pentru acetilcolina. Astfel, se poate aprecia ca celor doua capete boselate de pe molecula de acetilcolina (-N (CH3)3, -O-CO-CH3) trebuie sa le corespunda doua adancituri pe situsul receptor, capatului cationic al acetilcolinei (-N (CH3)3) trebuie sa ii corespunda pe situsul receptor un sediu anionic, iar legaturii esterice a moleculei de acetilcolina trebuie sa ii corespunda pe situsul receptor un sediu esterofil.
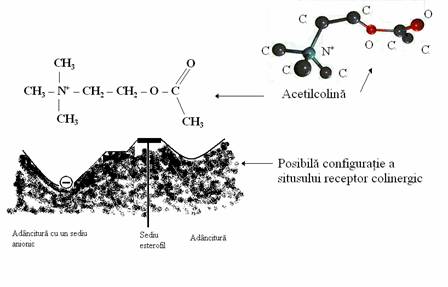
Fig. nr. 7.2. Posibila configuratie a situsului receptor colinergic plecand de la structura chimica a acetilcolinei. Celor doua capete boselate ale acetilcolinei ar trebui sa le corespunda doua adancituri la nivelul situsului receptor, capatului cationic al acetilcolinei ar trebui sa ii corespunda un sediu anionic, iar legaturii esterice a acetilcolinei ar trebui sa ii corespunda un sediu esterofil.
Cercetarea efectelor diferitelor substante agoniste sau antagoniste ale receptorilor colinergici a aratat insa ca populatia receptorilor colinergici nu este omogena in ceea ce priveste structura sterica a situsului receptor, existand probabil importante diferente intre receptorii colinergici din acest punct de vedere. Astfel, alcaloidul muscarina continut in ciuperca Amanita muscaria, reproduce numai anumite efecte ale acetilcolinei, pe cand alcaloidul nicotina, continut in planta Nicotiana tabacum (planta de tutun) reproduce alte efecte ale acetilcolinei, diferite de cele reproduse de muscarina. Este astfel evident ca receptorii colinergici asupra carora actioneaza muscarina sunt diferiti, in ceea ce priveste structura sterica a situsului receptor, de receptorii asupra carora actioneza nicotina. Receptorii asupra carora actioneaza nicotina au fost numiti receptori colinergici de tip nicotinic sau receptori nicotinici si notati cu N, pe cand receptorii asupra carora actioneaza muscarina au fost numiti receptori colinergici de tip muscarinic sau receptori muscarinici si notati cu M.
Receptorii nicotinici, plecand de la efectele pe care le prezinta nicotina, sunt situati in sistemul nervos central si in majoritatea sinapselor priferice ale neuronilor situati in sistemul nervos central si care trimit terminatii axonale in periferie cum sunt sinapsele neuromusculare (placile motorii) ale fibrelor motorii somatice, sinapsele ganglionare ale fibrelor preganglionare atat simpatice cat si parasimpatice, sinapsele fibrelor care inerveaza glanda medulosuprarenala, considerata un mare ganglion simpatic modificat. Studierea diverselor substante active asupra receptorilor nicotinici a aratat ca nici populatia acestor receptori nu este omogena. Exista o substanta numita hexametoniu, care este activa numai la nivelul receptorilor nicotinici situati la nivelul ganglionilor vegetativi, atat simpatici cat si parasimpatici, si asupra receptorilor nicotinici situati la nivelul glandei medulosuprarenale, dar nu asupra receptorilor nicotinici situati la nivelul placii motorii. In schimb exista o alta substanta numita decametoniu care nu este activa asupra receptorilor nicotinici sensibili la hexametoniu, dar este activa asupra receptorilor nicotinici situati la nivelul placii motorii, pe care nu actioneaza hexametoniul. Deci exista cel putin doua tipuri de receptori nicotinici, unii situati la nivelul ganglionilor vegetativi, atat simpatici cat si parasimpatici si la nivelul glandei medulosuprarenale, numiti receptori nicotinici ganglionari notati cu NN (indicele N provine de la nodulus = ganglion) si altii situati la nivelul sinapselor neuromusculare somatice (placilor motorii), numiti receptori nicotinici musculari notati cu NM (indicele M provenind de la musculus muschi). Din analiza structurii chmice a celor doua substante se pot trage concluzii privind structura situsurilor receptoare ale celor doua tipuri de receptori nicotinici. Faptul ca cele doua substante au doua capete cationice sugereaza ca pe receptorul nicotinic exista doua sedii anionice. La nivelul receptorilor NN de la nivelul ganglionilor, unde actioneaza hexametoniul, distanta intre cele doua sedii anionice trebuie sa fie aproximativ egala cu lungimea unei catene de 6 atomi de carbon pe cand la nivelul receptorilor NM de la placa motorie, unde actioneaza decametoniul, distanta intre sediile anionice trebuie sa fie aproximativ egala cu lungimea unei catene de 10 atomi de carbon. Acetilcolina avand un singur capat cationic rezulta ca pentru actionarea unui receptor nicotinic sunt necesare concomitent 2 molecule de acetilcolina:
|
H3C-CO-O
![]()
(CH3)3N+ CH2-CH2 Acetilcolina
(CH3)3N - (CH2 CH2 CH2 CH2 CH2 CH2) - N+(CH3)3 Hexametoniu
(CH3)3N - (CH2 CH2 CH2 CH2 CH2 CH2 CH2 CH2 CH2 CH2) - N+(CH3)3 Decametoniu
![]() CH2- CH2 - N CH3)3
Acetilcolina
CH2- CH2 - N CH3)3
Acetilcolina
O-CO-CH3
Fig. nr.7.3. Structura chimica comparativa a acetilcolinei, hexametoniului si decametoniului. Faptul ca atat hexametoniul cat si decametoniul au 2 capete cationice sugereaza ca la nivelul receptorului nicotinic exista 2 sedii anionice ceea ce presupune ca pentru actionarea unui astfel de receptor sunt necesare 2 molecule de acetilcolina. Distanta intre cele doua sedii anionice este egala cu lungimea unei catene de 6 atomi de carbon in cazul receptorilor NN (ganglionari) pe care se fixeaza hexametoniul si de 10 atomi de carbon in cazul receptorilor NM (musculari) pe care se fixeaza decametoniul.
Prin clonare s-a putut cunoaste cu mare exactitate structura receptorului nicotinic. Acesta este un canal ionic pentru sodiu, receptor dependent. Structura sa este foarte asemanatoare cu structura receptorilor GABA-ergici de tip GABAA, receptorilor glicinergici si receptorilor serotoninergici de tip 5-HT3, dar structura receptorului nicotinic a fost cel mai bine studiata. El este format din 5 subunitati polipeptidice care pot fi de mai multe tipuri notate cu , , , si . Fiecare din aceste subunitati este constituita din patru segmente transmembranare numite motive si notate cu M1-4 numerotarea facandu-se dinspre capatul amio-terminal catre capatul carboxi-terminal. Ambele capete sunt situate la nivelul suprafetei externe a membranei celulare dar capatul amino-terminal este mai lung decat capatul carboxiterminal. Exista de asemenea doua anse intracelulare, care unesc segmentele M1 si M2 si, respectiv, M3 si M4, precum si o ansa extracelulara care uneste segmentele M2 si M3. Motivul M2 are o conformatie helicoidala si participa la constituirea porului ionic. Cinci astfel de subunitati se asociaza pentru constituirea unui canal ionic de asa natura incat motivele lor M2 constituie porul ionic al canalului.
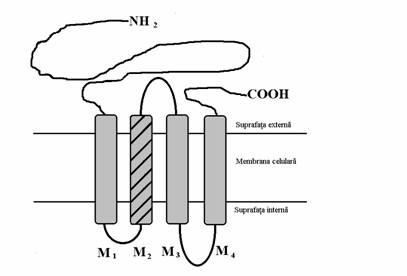
Fig. nr.7.4. Reprezentarea schematica a structurii subunitatii a receptorului nicotinic. Capatul carboxi-teminal, ansa care uneste motivele M2 si M3 si capatul amino-terminal care este lung fiind situate extracelular fac portiunea extracelulara a subunitatii sa fie mai voluminoasa decat portiunea sa intracelulara. Motivul M2 are o dispozitie helicoidala si contribuie la constituirea porului ionic.
Capatul extracelular al acestor subunitati fiind mai voluminos decat capatul intracelular, receptorul prezinta un fel de boselura extracelulara avand probabil o forma asemanatoare cu o floare de crin sau de lalea cu cinci petale. Pe subunitatea exista situsul receptor pentru acetilcolina si la constituirea unui canal ionic participa intotdeauna doua subunitati ceea ce face ca la nivelul receptorului nicotinic sa existe doua situsuri de legare pentru acetilcolina. Celelalte trei subunitati care participa la constituirea canalului ionic pot fi diferite. La nivelul fibrei musculare striate receptorul nicotinic este format din doua subunitati si cate o subunitate , si dar cel putin in unele fibre musculare mature subunitatea este inlocuita cu o subunitate . In sistemul nervos receptorii nicotinici se pot constitui din multiple alte combinatii pentamerice, dar probabil ca intotdeauna sunt prezente doua subunitati . A fost descrisa spre exemplu combinatia 23. In plus de aceasta exista cel putin 9 tipuri de subunitati notate cu 1-9 si cel putin 4 subunitati notate cu 1-4. Mai mult decat atat, in sistemul nervos central au fost descrisi receptori nicotinici presinaptici precum si receptori nicotinici care sunt canale de calciu, nu canale de sodiu. Aceasta face probabila existenta a mult mai multor tipuri de receptori nicotinici decat cele doua tipuri clasice, NN si NM, descrise anterior, dar pana la ora actuala nu se cunosc liganzi specifici decat in sensul existentei acestor doua tipuri.
Fixarea concomitenta a doua molecule de acetilcolina de un receptor nicotinic deschide canalul ionic respectiv si activeaza curentul de sodiu care depolarizeaza membrana celulara care il contine. Fluxul ionilor de sodiu prin canal este foarte intens, de aproximativ 5 107 ioni pe secunda, ceea ce are drept consecinta aparitia unui potential postsinaptic excitator. Daca se deschid suficient de multe canale ionice, potentialul postsinaptic excitator este suficient de amplu pentru a genera un potential propagat. Aceasta depinde de cantitatea de acetilcolina eliberata in fanta sinaptica si de densitatea receptorilor nicotinici in membrana respectiva. La nivelul placii motorii spre exemplu densitatea receptorilor nicotinici poate atinge 10 000 receptori m2.
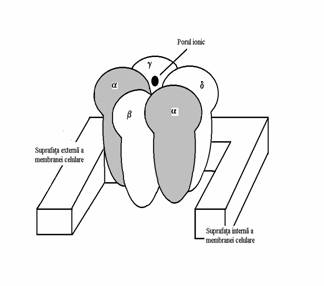
Fig.nr.7.5. Reprezentarea schematica a structurii receptorului nicotinic. Cinci subunitati, dintre care doua subunitati care contin situsul receptor pentru acetilcolina, se asociaza pentru a delimita un canal ionic. Capatul extracelular al receptorului este de dimensiuni mai mari decat capatul intracelular. Fixarea a doua molecule de acetilcolina pe cele doua subunitati determina o asemenea modificare a conformatiei sterice a receptorului incat canalul de sodiu se deschide permitand trecerea ionilor de sodiu prin canal sub influenta gradientului electrochimic si activarea curentului de sodiu care va depolariza membrana care il contine.
Receptorii muscarinici, spre deosebire de receptorii nicotinici, fac parte din categoria receptorilor cuplati cu proteinele G. Prin clonare s-a putut cunoaste suficient de clar structura acestor receptori. Se stie astazi ca exista cinci gene care codifica cel putin cinci tipuri distincte de receptori muscarinici notate cu m1-5. Toate cele cinci tipuri sunt constituite dintr-un lant polipeptidic care prezinta sapte segmente transmembranare unite prin trei anse intracelulare si trei anse extracelulare si prezentand un capat amino-terminal extracelular si un capat carboxi-terminal intracelular. Capatul amino-terminal impreuna cu cele trei anse extracelulare sunt uneori numite segmente extracelulare ale receptorului notate cu o1-4, segmentul o1 fiind capatul amino-terminal, iar capatul carboxi-terminal impreuna cu cele trei anse intracelulare sunt numite uneori segmente intracelulare ale receptorului notate cu i1-4, segmentul i4 fiind capatul carboxi-terminal. Segmentele transmembranare contin 20-25 aminoacizi hidrofobi si sunt practic identice pentru toti receptorii muscarinici precum si pentru receptorul adrenergic de tip 2 si receptorul pentru rodopsina, toti acesti receptori constituindu-se intr-o aceeasi familie de receptori farmacologici. Ansa a treia intracelulara (segmentul i3 al receptorului) este foarte lunga reprezentand aproximativ 25% din intreaga lungime a receptorului si este unica pentru fiecare subtip de receptor. Totusi primii 18 aminoacizi ai acestei anse se constituie in aceeasi succesiune pentru receptorii m1, m3 si m5 diferita de succesiunea care exista la receptorii m2 si m4. Ansa a treia intracelulara este cea prin intermediul careia receptorii se cupleaza cu proteine G. Receptorii m1, m3 si m5 se cupleaza cu o proteina Gs, pe cand receptorii m2 si m4 se cupleaza cu o proteina Gi. A fost insa demonstrat ca un acelasi tip de receptori se poate cupla cu mai mult decat o singura proteina G. Astfel unii din receptorii muscarinici se pot cupla de asemenea cu o proteina Gq.
Plecand de la astfel de considerente se poate aprecia ca fixarea acetilcolinei de receptorii muscarinici determina, in functie de tipul de receptor actionat, fie o crestere a cantitatii de AMPc intracelular prin stimularea adenilatciclazei, daca receptorul este cuplat cu o proteina Gs, fie o scadere a cantitatii de AMPc intracelular, daca receptorul este cuplat cu o proteina Gi, fie o crestere a formarii de diacilglicerol si inozitoltrifosfat (IP3), acesta din urma determinand eliberarea calciului din depozitele intracelulare si cresterea concentratiei intracelulare a calciului, daca receptorul se cupleaza cu o proteina Gq. Receptorii cuplati cu proteine Gs si Gq vor avea in special efecte stimulante pe cand receptorii cuplati cu proteina Gi vor avea efecte inhibitorii. In plus, receptorii cuplati cu proteina Gi determina de asemenea o crestere a intensitatii curentului de potasiu care este un curent inhibitor. Aceasta crestere a curentului de potasiu se poate explica prin deschiderea canalelor de potasiu, fie de catre subunitatea a proteinei Gi desprinsa de subunitatile , fie de subunitatile ale acestei proteine desprinse de subunitatea , fie de cresterea cantitatii de ATP ca urmare a inhibarii adenilatciclazei prin activarea proteinei Gi, adenilatciclaza fiind enzima care transforma ATP in AMPc. Unul sau altul dintre cele trei mecanisme intervin cu o pondere diferita la diferite specii animale dar intotdeauna stimularea vagala determina o crestere a intensitatii curentului de potasiu la nivelul cordului produsa de acetilcolina eliberata ca urmare a acestei stimulari, prin intermediul unor receptori muscarinici de tip m2.
Desi nu exista inca agonisti selectivi pentru cele 5 subtipuri de receptori muscarinici, exista totusi posibilitatea manipularii lor selective din punct de vedere farmacologic deoarece pentru 4 din cele 5 tipuri de receptori muscarinici exista antagonisti selectivi. Din aceste considerente se accepta notarea acestor receptori cu majuscule, respeciv ei pot fi notati M1-5. Antagonistii selectivi pentru receptorii M1-4 sunt, in ordine, pirenzepina, tripitramina, darifenacina si substanta al carei nume de cod este PD102807.
Efectele acetilcolinei asupra organismului depind de distributia diverselor tipuri de receptori colinergici in diferite aparate si sisteme. La nivelul sistemului nervos central, in diverse structuri, exista toate tipurile de receptori colinergici, atat nicotinici cat si muscarinici. In periferie receptorii nicotinici de tip NN se gasesc cu precadere la nivelul sinapselor din ganglionii vegetativi, atat simpatici cat si parasimpatici, si la nivelul glandei medulosuprarenale. Stimularea receptorilor ganglionilor parasimpatici depolarizeaza neuronii ganglionari parasimpatici iar impulsul nervos astfel initiat va deretmina eliberarea de acetilcolina in sinapsele parasimpatice terminale. Stimularea receptorilor ganglionilor simpatici depolarizeaza neuronii ganglionari simpatici iar impulsul nervos astfel initiat va deretmina eliberarea de noradrenalina in sinapsele simpatice terminale. Stimularea receptorilor nicotinici de la nivelul glandei medulosuprarenale determina eliberarea de adrenalina si noradrenalina in circulatia sanguina. Receptorii nicotinici de tip NM se gasesc la nivelul sinapsei neuromusculare somatice (placii motorii) si stimularea lor determina contractia musculaturii striate.
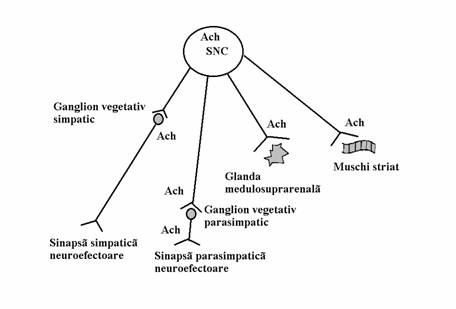
Fig.nr.7.6. Schema inervatiei colinergice. Acetilcolina este neurotransmitator in sistemul nervos central, la nivelul sinapselor periferice ale axonilor neuronilor situati in sistemul nervos central si la nivelul sinapselor periferice ale axonilor neuronilor situati in ganglionii vegetativi parasimpatici. Sistemul nervos simpatic a fost reprezentat printr-o fibra preganglionara scurta si o fibra postganglionara lunga iar sistemul nervos parasimpatic a fost reprezentat printr-o fibra preganglionara lunga si o fibra postganglionara scurta. SNC = sistem nervos central. Ach = acetilcolina
Receptorii muscarinici se gasesc cu precadere la nivelul sinapselor parasimpatice neuroefectorii, desi exista si unele sinapse neuroefectorii simpatice care contin receptori muscarinici. Spre deosebire de receptorii nicotinici periferici a caror stimulare produce intotdeauna declansarea unei activitati (depolarizare a neuronului ganglionar, contractia musculaturii striate sau secretia de adrenalina si noradrenalina) stimularea receptorilor muscarinici periferici foarte adesea moduleaza intensitatea activitatii unor viscere. In functie de densitatea unora sau altora dintre receptorii muscarinici in diverse structuri, efectele acetilcolinei pot fi stimulante ale activitatii sau deprimante ale acesteia. La nivelul tubului digestiv acetilcolina are in general efecte stimulante. Ea determina cresterea secretiilor digestive, inclusiv de acid clorhidric si pepsina la nivelul stomacului, favorizeaza golirea stomacului, crescand peristaltica gastrica si relaxand pilorul, accelereaza tranzitul intestinal, accelerand miscarile peristaltice si relaxand sfincterele. Tot efecte stimulante prezinta acetilcolina asupra bronhiilor, producand bronhoconstrictie si cresterea secretiei traheobronsice, asupra ureterelor, carora le creste peristaltica, si asupra organelor cavitare, vezica biliara si vezica urinara, carora le favorizeaza evacuarea continutului prin contractia musculaturii vezicale si relaxarea sfincterelor. Exista de asemenea receptori muscarinici la sinapsele simpatice de la nivelul glandelor sudoripare prin intermediul carora acetilcolina determina cresterea secretiei sudorale. Asupra aparatului cardio-vascular acetilcolina are in general efecte deprimante. Ea scade frecventa cordului, scade excitabilitatea, scade viteza de conducere atrio-ventriculara si scade contractilitatea fibrei miocardice. Aceste efecte sunt prezente insa numai la nivelul atriilor si portiunii superioare a jonctiunii atrio-ventriculare, ventriculii nefiind inervati vagal (parasimpatic). Asupra vaselor sanguine acetilcolina produce vasodilatatie, dar acest efect este datorat eliberarii de monoxid de azot (NO) de la nivelul celulelor endoteliale, probabil prin stimularea monoxid de azot sintetazei endoteliale (eNOS). Asupra ochiului stimularea receptorilor muscarinici determina contractia musculaturii circulare a irisului cu producerea de mioza, contractia musculaturii ciliare cu bombarea cristalinului si fixarea acestuia pentru privirea de aproape, scaderea presiunii intraoculare prin favorizarea resorbtiei umorii apoase, cresterea secretiei lacrimale.
Exista foarte multe modalitati de interferare farmacologica a sistemului colinergic. Aceasta influentare se poate face prin actionarea directa a receptorilor colinergici cu medicamente agoniste sau blocante (antagoniste) ale acestor receptori, sau prin actionarea lor indirecta, prin intermediul acetilcolinei endogene.
In ceea ce priveste actionarea directa a receptorilor colinergici, existenta mai multor tipuri de receptori colinergici permite actionarea lor selectiva, diferentiata. Agonistii receptorilor muscarinici produce in general efecte asemanatoare cresterii tonusului sistemului nervos parasimpatic, motiv pentru care aceste substante se numesc de obicei substante parasimpatomimetice sau, uneori, vagomimetice, plecand de la considerentul ca principalul nerv parasimpatic al organismului este nervul vag. Antagonistii sau blocantii receptorilor muscarinici produc in general efecte asemanatoare cu diminuarea sau inlaturarea tonusului parasimpatic, motiv pentru care se numesc de obicei substante parasimpatolitice sau, uneori, vagolitice. Agonistii receptorilor nicotinici produc efecte tipice pentru nicotina si sunt numiti de obicei substante nicotinice. Blocantele receptorilor nicotinici de tip NN produc practic o paralizie chimica a ganglionilor vegetativi si din acest motiv se numesc substante ganglioplegice. In fine, substantele care actioneaza selectiv asupra receptorilor NM se numesc prin traditie substante curarizante.
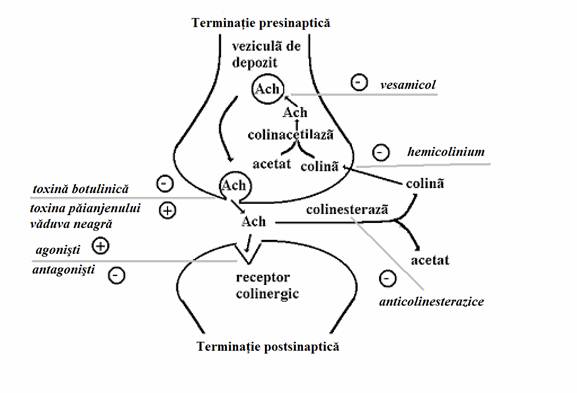
Fig.nr.7.7. Principalele modalitati de influentare farmacologica a sinapsei colinergice. Ach = aceticolina, + = stimuleaza procesul, - = inhiba procesul
Actionarea indirecta a receptorilor colinergici se poate face prin cresterea sau scaderea disponibilului de acetilcolina in fanta sinaptica. Cresterea disponibilului de acetilcolina in fanta sinaptica va creste activitatea sinapsei respective pe cand scaderea disponibilului acesteia va antrena o diminuare a activitatii sinapsei care poate merge pana la paralizia acesteia. Practic exista foarte multe posibilitati in acest sens dar nu toate au implicatii terapeutice reale. In ordinea fireasca a desfasurarii evenimentelor la nivelul sinapsei, o prima etapa in care se poate interveni consta in blocarea canalelor de sodiu ale terminatiei presinaptice prin tetrodotoxina, toxina care se gaseste in organele interne ale pestelui balon sau fugu, o delicatesa culinara pentru japonezi. Aceasta toxina foarte puternica impiedica transmiterea influxului nervos catre sinapse producand paralizie cu moarte care survine de obicei prin paralizia muschilor respiratori. Intr-o a doua etapa exista posibilitatea blocarii canalelor de calciu presinaptice cu impiedicarea influxului de calciu care sa elibereze acetilcolina in fanta sinaptica. Acest fenomen poate fi produs uneori de doze mari de antibiotice aminoglicozidice care, prin acest mecanism, pot determina paralizia muschilor respiratori, mai ales daca aminoglicozidul a fost administrat in doze mari intraoperator. De asemenea, prin acest mecanism antibioticele aminoglicozidice pot potenta medicamentele curarizante si pot agrava miastenia gravis. Impiedicarea eliberarii de acetilcolina in fanta sinaptica este posibila de asemenea prin toxina botulinica dar in acest caz toxina actioneaza asupra proteinelor implicate in procesul de eliberare a acetilcolinei (a se vedea 6. Influentarea farmacologica a sistemului nervos). Prin acest mecanism toxina botulinica produce paralizia sinapselor colinergice. In doze mari moartea se produce prin paralizia muschilor respiratori. In doze mici, administrate local, toxina botulinica poate fi insa utila terapeutic. Se poate utiliza pentru tratamentul unor contracturi patologice ale musculaturii striate, cum se intampla spre exemplu in torticolis sau in parezele spastice, in combaterea unor stari de hipersecretie sudorala, cum se intampla in dishidroza, sau in cosmetica pentru combaterea ridurilor. Tot asupra proteinelor implicate in procesul de eliberare a acetilcolinei in fanta sinaptica intervine toxina paianjenului numit vaduva neagra (black widow spider). Spre deosebire de toxina botulinica toxina acestui paianjen produce insa o descarcare masiva de acetilcolina in fanta sinaptica. La nivelul musculaturii striate se produc initial spasme musculare generalizate iar ulterior apare o paralizie generalizata, moartea survenind de asemenea prin paralizia muschilor respiratori. Exista si posibilitatea impiedicarii recaptarii, fie a colinei din fanta sinaptica in terminatia presinaptica prin hemicolinium, fie a acetilcolinei din citoplasma terminatiei presinaptice in granula de depozit, prin vesamicol. Ambele substante determina o paralizie a sinapselor colionergice prin epuizarea depozitelor presinaptice de acetilcolilna. In fine, exista posibilitatea cresterii disponibilului de acetilcolina in fanta sinaptica prin inhibarea metabolizarii acesteia de catre colinesteraza. Substantele care inhiba activitatea colinesterazei sunt cunoscute sub numele de substante anicolinesterazice si prezinta un mare interes atat terapeutic cat si toxicologic.
| Contact |- ia legatura cu noi -| | |
| Adauga document |- pune-ti documente online -| | |
| Termeni & conditii de utilizare |- politica de cookies si de confidentialitate -| | |
| Copyright © |- 2025 - Toate drepturile rezervate -| |
|
|
|||
|
|||
Analize pe aceeasi tema | |||
|
| |||
|
|||
|
|
|||