|






 |  |

| Biologie | Botanica | Chimie | Didactica | Fizica | Geografie |
| Gradinita | Literatura | Matematica |
Chimie
|
|
Qdidactic » didactica & scoala » chimie Celuloza - starea naturala. Definitia, biosinteza celulozei, structura chimica a celulozei, structura moleculara |
Celuloza - starea naturala. Definitia, biosinteza celulozei, structura chimica a celulozei, structura moleculara
Celuloza
1. Starea naturala. Definitia
Celuloza este polimerul natural cel mai raspindit. Este constituentul de baza al tesuturilor vegetale si componenta principala a membranei celulare din toate plantele superioare, inclusiv din speciile lemnoase.
In tesuturile vegetale celuloza se afla sub forma de fibre, avind rolul unei substante schelet, de care sint fixati toti ceilalti componenti chimici principali si secundari ai lemnului cum sint: lignina, hemicelulozele si substantele extractibile. Cu aceste substante (substante incrustante) celuloza se distribuie simplu in amestec sau este intim asociata.
Practic este aproape imposibil a separa celuloza chimic pura din peretele membranei celulare vegetale fara o deteriorare mai mult sau mai putin avansata a structurii sale chimice. Este necesar de a face aceasta precizare, pentru a deosebi celuloza chimic pura, sau celuloza nativa de celuloza tehnica care se obtine prin dezincrustarea sau fierberea lemnului. Aceasta reprezinta de fapt un amestec de celuloza chimica si diversi alti componenti cu care ea se asociaza in mod natural in plante. Celuloza din lemn are o fibra mai scurta decit celuloza din bumbac. Astfel, fibrele de celuloza din lemn de molid au lungimea de 2,6 - 3,8 nm, in timp ce fibrele de celuloza din lemnul de fag au o lungime cuprinsa intre 0,7 - 1,7 nm. Lungimea fibrelor de celuloza din bumbac ajunge pina la 40 - 50 nm. Din acest motiv, in cele ce urmeaza se va studia doar celuloza chimica propriu-zisa care se va numi simplu celuloza.
Din punct de vedere chimic, prin notiunea de celuloza se intelege o polizaharida naturala cu structura fibrilara, care rezulta prin condensarea unui numar variabil de unitati de D-glucoza, cu compozitia (C6H10O5)n.
Asadar, termenul de celuloza nu defineste o substanta chimica unitara, ci un agregat supramolecular format din mai multe catene macromoleculare de glucan, diferite ca lungime si asociate intre ele.
Continutul mediu de celuloza in plante este in general diferit. Singura varietate de celuloza naturala aproape pura o constituie bumbacul, care contine 91 - 93 % celuloza, in timp ce la celelalte plante continutul de celuloza este mai mic: in - 82,5 %, cinepa - 77,7 %, rafie - 78,7 %, iuta - 64,2 %, speciile lemnoase - 57 - 45 %, stuf - 48,7 %. La arbori continutul de celuloza variaza in limite destul de largi in functie de specia lemnoasa, zona de prelevare a probei, zona geografica, conditiile pedologice si mai putin in functie de inaltime, virsta si altitudine.
Tabelul 1
Valorile maxime in ordine descrescatoare ale continutului de celuloza din lemnul unor specii de arbori
|
Nr. Crt. |
Specia lemnoasa |
Continutul de celuloza, % |
Nr. Crt. |
Specia lemnoasa |
Continutul de celuloza, % |
|
1. |
Molid |
57,00 |
8. |
Ulm |
49,63 |
|
|
Brad |
55,09 |
9. |
Fag |
49,23 |
|
3. |
Pin |
55,07 |
10. |
Salcim |
48,72 |
|
4. |
Tei |
55,74 |
11. |
Mesteacan |
48,45 |
|
5. |
Salcie |
53,66 |
1 |
Carpen |
47,83 |
|
6. |
Paltin |
53,02 |
13. |
Stejar |
45,08 |
|
7. |
Plop |
52,56 |
|
|
|
Continutul cel mai mare de celuloza se intilneste la speciile de conifere: molid, brad si pin. Speciile de foioase au un continut mai redus de celuloza, valori maxime inregistrindu-se la tei, salcie si paltin si valori minime la mesteacan, carpen si stejar.
Deoarece unele dintre foioase, cum ar fi plopul, ulmul, fagul si mesteacanul contin cantitati apreciabile de celuloza si produc o cantitate anuala de masa lemnoasa mai mare, ele sint valorificate in mod prioritar pentru fabricarea celulozei si hirtiei.
Biosinteza celulozei
Celuloza se formeaza in plante in timpul procesului de fotosinteza, din CO2 si H2O, sub influenta luminii si in prezenta clorofilei. Din aceste substante prin ciclul stationar al fotosintezei se ajunge la unitati elementare de D-glucoza, capabile sa formeze lanturi macromoleculare de celuloza.
Conform uneia din teoriile existente, formarea polizaharidelor in plante urmeaza un mecanism enzimatic, in doua directii principale sub actiunea specifica a diferitelor enzime.
Intr-o prima etapa moleculele de D-glucoza se unesc succesiv sub actiunea enzimelor de tipul fosforilazei, care favorizeaza aparitia legaturilor C1 - C4 b-glicozidice dintre unitatile elementare de glucoza.
In cea de a doua etapa, enzima respectiva catalizeaza unirea simultana a unor fragmente de macromolecule realizate anterior, proces numit si bloc copolimerizare.
Se mai cere insa de explicat modul in care enzimele catalizeaza polimerizarea si cristalizarea simultana a lanturilor macromoleculare de celuloza intre ele. Teoretic exista doua variante, care schematic sint prezentate in fig. 1. Intr-o prima varianta redata grafic in fig. 1a, fibra elementara de celuloza se dezvolta dintr-o singura particula enzimatica cu forma sferica, avind diametrul de 8 nm, care prin extrudere intr-o anumita directie se alungeste devenind mai subtire pina la grosimea de 3,5 nm.

Aceasta ipoteza este in concordanta cu diametrul constant real al fibrei elementare, dar nu da nici o informatie in privinta continutului de celuloza paracristalina si amorfa din fibra de celuloza. Mai mult, ea nu explica o anumita lungime a fibrilei elementare de celuloza.
In cea dea doua varianta (fig. 1b si 1c) se considera ca sinteza fibrilei elementare de celuloza are loc din mai multe particule enzimatice, dezvoltarea acestora avind loc in directie laterala. La punctele de contact cu enzima, reteaua lanturilor macromoleculare de celuloza este construita intr-o maniera mult mai ordonata (paralele), constituind astfel asa-numita zona cristalina, in timp ce intre particulele enzimatice reteaua este mult mai putin ordonata, constituind zona amorfa. Pina in prezent, insa, sistemul si modalitatea de formare a fibrilelor elementare de celuloza nu au fost pe deplin elucidate.
Din studiul structurii si al formarii peretilor membranei celulare vegetale se pot trage totusi unele concluzii de baza:
Celuloza apare initial in peretele primar alaturi de hemiceluloze, pectine si lignina, servind drept cadru pentru sustinerea acestora. In timpul cresterii plantelor, celuloza se constituie sub forma unor microfibrile cu diametrul de 10 - 30 nm ingrosate in planul peretelui primar.
In peretele secundar, insa, microfibrilele de celuloza prezinta o orientare strict paralela si cu dimensiuni oarecum modificate, ceea ce denota ca procesul de biosinteza este influentat aici de alti factori decit in cazul peretelui primar.
Procesul final de formare a peretelui celular implica si formarea peretelui tertiar, care difera ca proprietati chimice si morfologice de ceilalti pereti ai membranei celulare vegetale.
2.3. Structura chimica a celulozei
Studiul structurii fibrelor celulozice se efectueaza cu mijloace adecvate la trei scari diferite:
a) la scara amicroscopica, cind se studiaza structura moleculara a celulozei. Aici se utilizeaza diferite cai indirecte, de regula chimice si fizico-chimice, se determina modul de legare a atomilor in cadrul macromoleculei, cit si a catenelor macromoleculare de celuloza in cadrul agregatului structural elementar (0,5 - 3 nm);
b) la scara submicroscopica, cind se studiaza structura fina sau supramoleculara. La aceasta scara exista posibilitatea de a observa direct dispunerea in cadrul fibrei de celuloza a elementelor specifice, numite fibrile (25 - 30 nm);
c) la scara microscopica, cind se studiaza structura microscopica a agregatului celulozic. Studiul la aceasta scara ofera posibilitatea observarii cu microscopul optic a detaliilor morfologice ale fibrei de celuloza (peste 500 nm).
2.3.1. Structura moleculara
Primele dovezi privind structura chimica a catenei macromoleculare de celuloza au fost furnizate de rezultatele procesului de hidroliza directa a acesteia cu acizi minerali concentrati. La hidroliza totala a celulozei cu acid clorhidric de 40 %, de exemplu, se formeaza cu randament de 96 - 98 % fata de cel teoretic D-glucoza. La hidroliza partiala a celulozei se formeaza dizaharida celobioza. Pe baza acestor rezultate s-a ajuns la concluzia ca unitatea structurala cea mai simpla, denumita si unitate elementara de structura (u.e.s.) a catenei macromoleculare de celuloza este anhidro-b-D-glucopiranoza
Prin reactiile de eterificare sau esterificare completa a celulozei se pot obtine sub forma de produsi finali numai eterii si respectiv esterii trisubstituiti ai celulozei. La hidroliza acestor produsi se obtin cantitati mai mari de trimetilglucoza si cantitati mici de tetrametilglucoza. De aici s-a ajuns la concluzia ca unitatea elementara de anhidro-b-D-glucopiranoza din celuloza contine trei grupe hidroxil alcoolice libere.
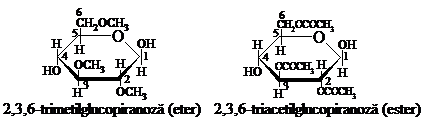
Din cele trei grupe hidroxil alcoolice ale unitatii elementare de anhidro-b-D-glucopiranoza, cea care se afla la atomul de C6 este grupa alcoolica primara, iar celelalte doua, care se afla la atomii de C2 si C3 sint grupe alcoolice secundare. Aceste grupe se deosebesc mult intre ele in ceea ce priveste reactivitatea. Astfel, grupele alcoolice primare sint de 5-6 ori mai reactive decit cele secundare.
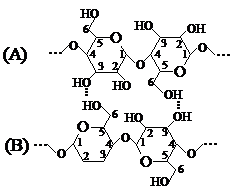
Unitatile elementare de anhidro-b-D-glucopiranoza din catena celulozei sint unite intre ele prin legaturi eterice specifice, denumite legaturi C1 - C4 b-glicozidice. Ele se realizeaza prin eliminarea unei molecule de apa de la hidroxilul glicozidic C1 al unei unitati de D-glucoza si hidroxilul alcoolic secundar de la C4 al unitatii de D-glucoza vecina. Atomul de oxigen prin care se leaga cele doua unitati elementare de structura (-O-) se numeste in acest caz oxigen glicozidic si se deosebeste de atomul de oxigen din puntea oxidica (C1-O-C5) care participa la formarea ciclului piranozic.
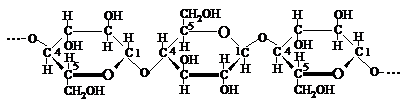
Unitatea elementara de anhidro-b-D-glucopiranoza se prezinta sub forma anomerului b, intrucit acest izomer intra in constitutia dizaharidei celobioza, care si este penultimul produs de hidroliza a celulozei.
Anume existenta legaturii C1-O-C4-glicozidice intre unitatile elementare ale catenei macromoleculare de celuloza determina stabilitatea ei scazuta la actiunea agentilor de hidroliza. Actiunea acestora, inclusiv a apei, la temperatura inalta provoaca desfacerea legaturilor glicozidice si depolimerizarea completa pina la D-glucoza.
Pentru a nu fi tensionat, ciclul piranozic poate avea doua conformatii spatiale, fie in forma de "baie", fie in forma de "scaun". Conformatia de tip "baie" este mai bogata in energie decit cea de tip "scaun". Prima are o stabilitate mai redusa si va fi mai rar intilnita in structura catenei macromoleculare de celuloza.
Pentru ciclul unitatii elementare de anhidro-b-D-glucopiranoza pot exista insa doua conformatii spatiale de tip scaun. Intr-o prima conformatie, denumita tip scaun C1, atomii de hidrogen sint in pozitie axiala (a), iar grupele hidroxil in pozitie ecuatoriala (e). In cea de a doua conformatie, denumita scaun 1C, atomii de hidrogen sint in pozitie ecuatoriala (e), iar grupele hidroxil in pozitie axiala (a).
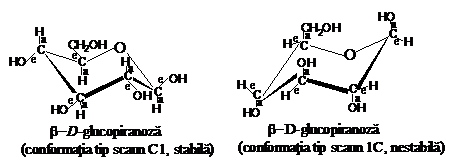
Pentru unitatea elementara de anhidro-b-D-glucopiranoza este mai stabila conformatia C1, care permite tuturor gruparilor substituente ( OH si CH2OH) asezarea pe legaturile ecuatoriale si, deci, o distantare maxima fata de ciclu.
Celuloza este considerata un compus macromolecular cu formula empirica (C6H10O5)n in care "n" este gradul de policondensare. Daca luam in consideratie ca o molecula de apa ramine legata de catena macromoleculara, formula celulozei mai poate fi prezentata: (C6H10O5)n H2O. Se ia in consideratie ca unitatea elementara de D glucopiranoza din extremitatea stinga a catenei are un atom de hidrogen in plus, iar cea din extremitatea dreapta o grupa hidroxil suplimentara:
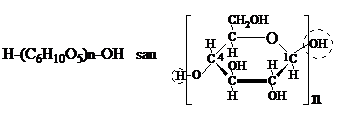
|
De mentionat ca celuloza prezinta anumite proprietati reducatoare (foarte neinsemnate).
In cazul compusilor macromoleculari naturali, respectiv a celulozei, unitatile elementare de anhidro-b-D-glucopiranoza sint capabile sa se roteasca unele fata de altele la 180O, ceea ce determina practic o indoire a catenei de celuloza din loc in loc. Datorita rotirii libere a ciclurilor piranozice de anhidro-b-D-glucopiranoza in ambele sensuri fata de atomul de oxigen glicozidic, catena macromoleculara de celuloza poate adapta, astfel, forme diferite. Pentru un nivel energetic aproape neschimbat, o catena macromoleculara de celuloza netensionata ar avea, comparativ cu una de amidon, o conformatie spatiala de tipul celei prezentate in fig.
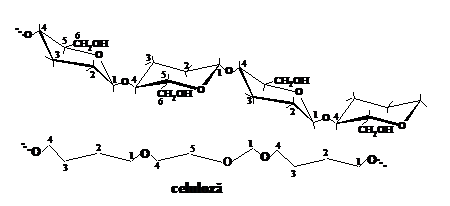
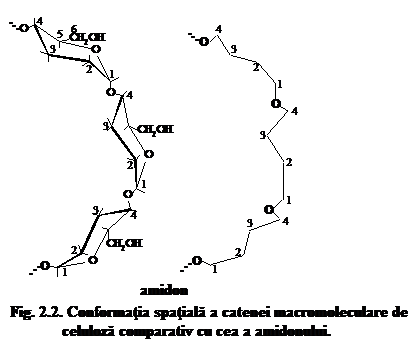
Daca admitem conformatia tip scaun C1 pentru unitatea elementara de anhidro-b-D-glucopiranoza, atunci conformatia spatiala a catenei macromoleculare de celuloza ar putea fi reprezentata sub forma unei panglici situata intre doua planuri imaginare (fig. 3) in care legaturile C1-O-C4 b-glicozidice ar alterna in forma de zig-zag desfasurat dupa un surub diagonal.
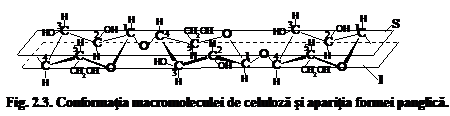
Planul superior imaginar al acestei panglici (S) este alcatuit succesiv din atomii C1, C3, C5 si C2, C4,O. Planul inferior imaginar al panglicii (I) este alcatuit si el succesiv din atomii C2, C4, O si C1, C3, C5. Asezarea succesiva a acelorasi atomi se repeta in catena de n/2 ori. Perpendicular pe planul superior si cel inferior al panglicii apar atomi de hidrogen in pozitie axiala, in timp ce grupele -OH si CH2OH sint situate in partile laterale ale panglicii, in pozitie ecuatoriala. De forma catenelor macromoleculare de celuloza depind o serie de proprietati fizice importante ale solutiilor sale, cum ar fi: viscozitatea, viteza de sedimentare, elasticitatea peliculelor de celuloza si a.m.d.
Forma ideala a catenei macromoleculare de celuloza, care confera rezistenta mecanica produselor obtinute din celuloza sau derivatilor acesteia, este cea de filament putin indoit.
Studiile mai aprofundate ale structurii catenei macromoleculare de celuloza au adus la ideea ca aceasta prezinta unele devieri de la formula ideala propusa pentru prima data de catre W. N. Haworth. Ca urmare, pentru a se ajunge la o formulare mai reala a structurii celulozei trebuie luate in consideratie unele completari si precizari, care au fost generate de o serie de date experimentale incontestabile.
Astfel, din cercetarile de hidroliza a celulozei din bumbac, care este cel mai pur material celulozic, s-a constatat ca aceasta este formata in proportie de 99 % din unitati de anhidro-b D-glucopiranoza. Diferenta de 1 % o reprezinta unitatile altor monozaharide. Cu alte cuvinte, catena macromoleculara de celuloza nu este alcatuita in exclusivitate numai din unitati de D-glucopiranoza, ci ea mai contine in cantitati foarte mici si resturi ale altor monozaharide. Prezenta resturilor de diferite monozaharide a fost temeinic dovedita si prin hidroliza totala a celulozei obtinuta din lemn. Astfel s-a constatat ca celuloza din molid contine pe linga D-glucoza cantitati apreciabile de manoza, iar cea din mesteacan - resturi de xiloza. In celuloza din specii de foioase s-au identificat si urme de arabinoza, manoza, galactoza si chiar acid glucuronic. Principalii insotitori ai celulozei din lemn sunt, deci, polizaharide mixte de tipul pentozanelor, hexozanelor si a acizilor poliuronici.
In procesul de biosinteza a celulozei, in afara de catenele macromoleculare de celuloza se formeaza si catene macromoleculare ale unor polizaharide mixte, in compozitia carora, pe linga resturi de anhidro-b D-glucopiranoza, intra si resturi ale altor monozaharide cu structura piranozica sau chiar furanozica. De exemplu, una dintre ele ar putea fi prezentata, avind in compozitie resturi de D-glucoza (G), D-xiloza (X), si de acid glucuronic (A.Gl.):
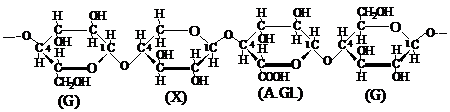
In consecinta, se poate afirma ca in diferitele materiale vegetale, catenele macromoleculare de celuloza nu au o compozitie chimica perfect identica. Preparatele celulozice tehnice, obtinute in diverse conditii si din anumite materii prime, se pot deosebi putin intre ele dupa compozitia chimica a resturilor de monozaharide care intra in molecula lor. Raportul dintre numarul unitatilor de glucoza si numarul unitatilor de alte monozaharide este diferit pentru diversele celuloze tehnice.
Tot din studiile de hidroliza a celulozei s-a mai constatat ca in catenele unor macromolecule de celuloza, un anumit numar de unitati elementare de D-glucoza se gasesc nu sub forma ciclica de D-glucopiranoza, ci sub forma deschisa, respectiv lineara, sau chiar sub forma ciclica de D-glucofuranoza.
Intr-o astfel de catena macromoleculara legatura intre unitatile elementare deschise sau aciclice si cele ciclice se realizeaza prin legaturi C1-O-C4 b-glicozidice obisnuite. Stabilitatea catenei macromoleculare de celuloza in ansamblul ei este asigurata in acest caz de existenta unor legaturi specifice C1 - C5 semiacetalice interne, care actioneaza in interiorul ciclului piranozic deschis intre C1 - C5:
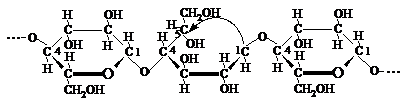
Fiind mult mai putin stabile la actiunea agentilor de hidroliza, legaturile C1 - C5 semiacetalice interne se vor rupe mai usor decit legaturile C1 - C4 b-glicozidice obisnuite, astfel scurtindu-se tot mai avansat catenele macromoleculare de celuloza.
Pornind de la ipoteza existentei unor unitati elementare de D-glucoza aciclice s-a ajuns la concluzia ca mentinerea in apropiere a catenelor macromoleculare se realizeaza nu numai prin legaturi de hidrogen sau prin legaturi Van der Waals, dar, in unele cazuri, si prin legaturi chimice mai deosebite, denumite in acest caz legaturi C1 - C5 semiacetalice laterale. Acestea actioneaza intre unitatile elementare de D-glucoza deschise, aflate de-a lungul unor catene macromoleculare de celuloza invecinate.
Aceste legaturi C1 - C5 semiacetalice laterale sint tot atit de putin stabile la actiunea agentilor de hidroliza ca si legaturile C1 - C5 semiacetalice interne. Ca urmare, prin hidroliza acida a celulozei se va produce mai intii ruperea legaturilor C1 - C5 semiacetalice laterale dintre catene, apoi a celor C1 - C5 semiacetalice interne din cadrul catenelor, urmate in final de desfacerea legaturilor C1 - C4 b-glicozidice normale dintre unitatile elementare de b-D-glucopiranoza.
In acest mod a putut fi explicata practic viteza mare a procesului de hidroliza a celulozei, ce are loc intr-o prima etapa pina la un grad mediu de polimerizare de cca. 850, dupa care, in cea de a doua etapa, ea decurge mult mai lent. Conform celor expuse, se pot concluziona doua aspecte importante:
Pe de o parte, pe anumite zone, de-a lungul catenelor macromoleculare de celuloza se pot distinge doua tipuri de legaturi intramoleculare sau intracatenare:
1. Legaturi C1 - C4 b-glicozidice intre unitatile elementare de D-glucopiranoza ciclice, analog formulei ideale propusa de Haworth;
2. Legaturi C1 - C5 semiacetalice interne in interiorul unitatilor elementare aciclice de D-glucoza.
Pe de alta parte, pe anumite zone, intre catenele macromoleculare de celuloza pot sa apara trei tipuri de legaturi intermoleculare sau intercatenare:
1. Legaturi de hidrogen intermoleculare.
2. Legaturi Van der Waals normale.
3. Legaturi C1 - C5 semiacetalice laterale.
Catenele macromoleculare de celuloza naturala se caracterizeaza printr-o asa-numita periodicitate stricta a structurii. Aceasta inseamna ca de-a lungul lor, dupa cca. 256 unitati elementare de D-glucopiranoza (X) urmeaza o unitate elementara de D-glucoza deschisa care genereaza aparitia legaturilor C1 - C5 semiacetalice interne in catene, sau a legaturilor C1 - C5 semiacetalice laterale intre catene. (fig. 4). Asemenea lanturi macromoleculare de celuloza se numesc catene primare (A) si au in general lungimi mici, respectiv grade de polimerizare de 64, 128, 256. Catenele macromoleculare de celuloza care se unesc intre ele prin legaturi C1 - C5 semiacetalice laterale formeaza catene secundare (B) si au lungimi mult mai mari. Cea mai mare parte a legaturilor intercatenare o reprezinta legaturile de hidrogen si Van der Waals normale.
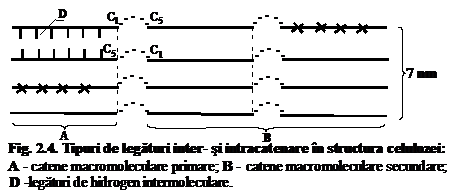
Din totalul legaturilor existente in celuloza circa 0,2 % se scindeaza de aproximativ 3000 ori mai repede decit legaturile C1 - C4 b-glicozidice normale. Aceste legaturi care se afla la distante bine determinate, respectiv dupa 465 30 unitati elementare de D-glucoza, au fost numite mai simplu legaturi slabe. Viteza reactiei de degradare a celulozei in mediu acid incepe sa se micsoreze simtitor cind gradul mediu de polimerizare al acesteia scade la 850 - 500, ceea ce inseamna ca sub aceasta valoare exista mai putine locuri slabe.
Asadar, referitor la structura moleculara a celulozei este important de retinut urmatoarele aspecte de baza:
1. Catenele macromoleculare din fibrilele de celuloza ale diferitelor materii prime vegetale nu sint constituite numai din resturi de anhidro-b-D-glucopiranoza. In compozitia lor pot intra in cantitati mici si resturi ale altor monozaharide, respectiv hexoze sau pentoze. Caracterul si numarul acestor resturi de monozaharide variaza de la un preparat celulozic la altul, fiind determinat de conditiile in care are loc biosinteza celulozei. Celuloza formata numai din resturi de anhidro-b-D-glucopiranoza se poate izola, probabil, in anumite conditii cu totul speciale.
2. Compozitia catenelor macromoleculare de celuloza separate din materialele vegetale nu este perfect identica. Diferitele preparate celulozice tehnice se pot deosebi intrucitva intre ele dupa compozitia chimica a unitatilor elementare de structura care intra in compozitia acestora si, deci, dupa stabilitatea lor fata de actiunea diferitilor agenti chimici.
3. In catenele macromoleculare de celuloza, in afara de unitatile elementare de D-glucoza, care au o structura ciclica piranozica, mai poate exista si un numar mic de resturi ale altor monozaharide ce poseda o structura ciclica furanozica, precum si unele unitati elementare de monozaharide cu o structura aciclica. Astfel de unitati elementare au o reactivitate chimica mult mai mare comparativ cu resturile de D-glucoza obisnuite.
4. Stabilitatea legaturilor C1 - C4 b-glicozidice din catenele macromoleculare de celuloza la actiunea diferitilor agenti chimici nu este constanta. Ea variaza in functie de caracterul si pozitia grupelor functionale din unitatea elementara de structura a monozaharidelor. In acelasi timp, legaturile C1 - C5 semiacetalice interne si laterale din si dintre resturile de D-glucoza deschise sint mult mai putin stabile la actiunea agentilor de hidroliza decit legaturile C1 - C4 b-glicozidice obisnuite dintre resturile de D-glucoza ciclice.
5. Structura moleculara reala a celulozei se abate sensibil de la formula ideala a lui W. N. Haworth, datorita transformarilor oxidative suferite de materialul fibros pe parcursul tratamentelor tehnologice de delignificare a lemnului.
3. Structura secundara
Prin structura secundara a celulozei, ca si a oricarui alt polimer, se intelege conformatia adoptata de catenele sale macromoleculare, sub influenta interactiunilor care apar si se manifesta intre lanturile invecinate. Din acest punct de vedere, trebuie retinut faptul ca, indiferent de lungimea lor, catenele macromoleculare de celuloza nu pot fi considerate ca individualizate ci, dimpotriva, ele se afla intr-o strinsa interdependenta. Aceasta interdependenta reciproca este determinata de aparitia unor legaturi de natura fizica sau chimica, care impreuna genereaza coeziunea de ansamblu a fibrei de celuloza.
Legaturile de natura chimica dintre catenele macromoleculare de celuloza se formeaza fie in procesul de biosinteza, fie la tratarea acesteia cu diferiti agenti chimici. In procesul de biosinteza a celulozei pot sa apara si sa se manifeste doua tipuri de legaturi chimice:
legaturi de tip esteric;
legaturi de tip C1 - C5 semiacetalice laterale.
Legaturile chimice de tip esteric apar in urma interactiei unei grupe carboxil, existenta din loc in loc la atomul C6 al unei unitati elementare de acid b-D-glucuronic din catena macromoleculara de celuloza (A) cu o grupa hidroxil alcoolica secundara libera de la atomul C2 dintr-o alta unitate elementara, aflata in catena macromoleculara de celuloza invecinata (B):
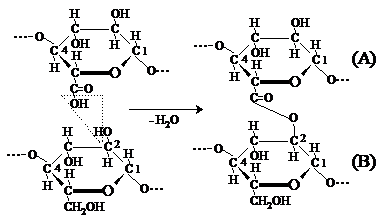
Legaturile de tip C1 - C5 semiacetalice laterale se formeaza intre unitatile elementare de D-glucoza sub forma deschisa (aciclica) aflate in doua catene invecinate.
Legaturile de natura fizica dintre catenele macromoleculare de celuloza pot fi:
legaturi de hidrogen;
legaturi de tip Van der Waals.
Legaturile de hidrogen se pot realiza intre doua grupe hidroxil apartinind celor doua unitati elementare de anhidro-b-D-glucopiranoza vecine, existente in aceeasi catena macromoleculara. Ele se numesc legaturi de hidrogen intramoleculare. Legaturile de hidrogen pot lua nastere insa si intre doua grupe hidroxil ce apartin unitatilor elementare din doua catene macromoleculare de celuloza vecine. In acest caz ele se numesc legaturi de hidrogen intermoleculare
Asadar, legaturile de hidrogen intra- si intermoleculare apar si se manifesta intre atomii de hidrogen care apartin grupelor alcoolice primare sau secundare si atomii de oxigen existenti in grupele hidroxil alcoolice de acelasi tip invecinate sau atomi de oxigen existenti in ciclul piranozic, fie in legatura C1 - C4 b-glicozidica.
Daca se admite conformatia C1 pentru unitatea elementara si conformatia tip panglica plana pentru catena macromoleculara de celuloza, atunci teoretic exista posibilitatea formarii urmatoarelor legaturi de hidrogen intramoleculare (fig. 5):
- intre atomul de oxigen din legatura glicozidica (O) C1 - C4 si atomul de hidrogen al gruparii -OH alcoolice primare de la atomul C6 din unitatea vecina din dreapta (A);
- intre acelasi atom de oxigen glicozidic si atomul de hidrogen din grupa -OH alcoolica secundara aflata la atomul C2 din unitatea vecina din stinga (B);
intre acelasi atom de oxigen glicozidic si atomul de hidrogen al gruparii -OH alcoolica secundara aflata la atomul C3 din unitatea vecina din dreapta (C);
- intre atomul de oxigen semiacetalic sau oxidic din ciclul piranozic si atomul de hidrogen al gruparii -OH alcoolica secundara aflata la atomul C3 din unitatea vecina din dreapta (D).

Cea mai convenabila pozitie sterica pentru a forma legaturi de hidrogen cu oxigenul glicozidic au gruparile alcoolice secundare de la C2 si C3. Legaturile de hidrogen care le formeaza ele cu acest oxigen se mai numesc legaturi de hidrogen helatice. Din schema ce urmeaza de asemenea se pot observa usor legaturile de hidrogen intramoleculare mentionate mai sus:
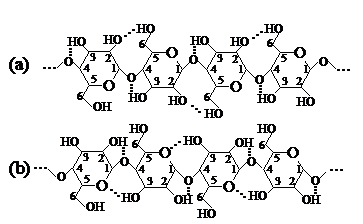
In schema (a) se observa posibilitatea aparitiei legaturilor de hidrogen intre hidrogenul grupei -OH alcoolica primare aflata la atomul C6 si oxigenul grupei -OH alcoolice secundare de la atomul C2 din unitatea vecina din stinga. O asemenea legatura de hidrogen intramoleculara este insa putin probabila, intrucit daca gruparea -OH alcoolica de la C6 ar fi blocata intr-un asemenea mod, intre catenele celulozei nu s-ar mai putea realiza practic si legaturi de hidrogen intermoleculare. Din acest motiv este mult mai posibila schema (b) unde se demonstreaza helatizarea grupei -OH de la atomul C2 cu oxigenul glicozidic. In acest caz gruparile -OH alcoolice primare de la C6 ramin disponibile pentru formarea legaturilor de hidrogen intermoleculare. Analizind natura legaturilor de hidrogen intramoleculare realizate de grupele -OH alcoolice secundare de la atomul C2 cu oxigenul glicozidic se constata ca ele sint niste legaturi directoare, care se opun intinderii catenei macromoleculare de celuloza. Legaturile de hidrogen formate de grupele -OH de la atomul C3 cu oxigenul din ciclul piranozic sint stabilizatoare si favorizeaza conformatia spatiala a catenei de celuloza sub forma de panglica plana.
O serie de proprietati fizico-chimice ale celulozei, cum ar fi: solubilitatea, umflarea, reactivitatea fata de anumiti agenti chimici s.a., sint insa in mare masura determinate nu atit de legaturile de hidrogen intramoleculare, cit, mai ales, de legaturile de hidrogen intermoleculare. Ele se realizeaza intre atomul de oxigen din grupa -OH alcoolica primara de la atomul C6 si atomul de hidrogen al gruparii -OH alcoolice secundare de la atomul C3 al unei alte catene macromoleculare vecina:
Este evident ca, cu cit numarul grupelor hidroxil libere va fi mai mare si cu cit catenele de celuloza vor fi mai apropiate intre ele, cu atit va creste posibilitatea formarii legaturilor de hidrogen intermoleculare.
Energia legaturilor de hidrogen din diversele materiale celulozice variaza de la 2-3 pina la 6-7 Kcal mol, iar distanta intre catene, mai precis intre atomii care formeaza legaturile de hidrogen (-OH OH) este de 0,267-0,275 nm.
Faptul ca grupa hidroxil de la atomul C3 din unitatile elementare de anhidro-b-D-glucopiranoza este solicitata atit la formarea legaturilor de hidrogen intramoleculare cit si intermoleculare demonstreaza ca aceasta are cea mai scazuta reactivitate.
Prezenta legaturilor de hidrogen intermoleculare conditioneaza capacitatea de patrundere a diversilor reactivi chimici in fibra de celuloza si, deci, limiteaza sau mareste posibilitatea de umflare, solubilizare si reactie a acesteia. Atunci cind patrunde o molecula de apa sau reactiv chimic prin partile laterale ale fibrei de celuloza si respectiv printre catenele macromoleculare, care sint dispuse paralel cu axa fibrei, are loc mai intii o indepartare relativa a unora fata de altele. Acest proces are ca efect final slabirea sau chiar ruperea legaturilor de hidrogen intermoleculare. In aceasta faza are loc o umflare puternica a fibrei de celuloza. Pe masura ce agentul chimic patrunde tot mai mult intre catene, are loc ruperea totala a legaturilor de hidrogen intermoleculare, ajungindu-se astfel in final la individualizarea tuturor catenelor macromoleculare. In acelasi timp pot sa apara si ruperi ale legaturilor de hidrogen intramoleculare, urmate de scindari ale legaturilor glicozidice cu o scurtare a catenei macromoleculare, adica cu o scadere a gradului de polimerizare.
Concluzionam ca legaturile de hidrogen intra- si intermoleculare din materialele celulozice determina intr-o masura apreciabila, atit pozitia reciproca a catenelor macromoleculare de celuloza, cit si proprietatile fizico-chimice ale acestora.
Legaturile de tip Van der Waals actioneaza atit intre grupele polare, cit si intre grupele nepolare ale catenelor macromoleculare de celuloza, contribuind si ele la coeziunea generala a fibrei de celuloza. Energia de interactiune a acestor legaturi este de 2-3 ori mai mica fata de cea a legaturilor de hidrogen, distanta minima de actiune dintre molecule fiind de 0,275 nm pina la 0,3 - 0,4 nm. Cu cresterea distantei dintre catenele macromoleculare de celuloza, intensitatea actiunii acestor forte scade brusc.
3.3. Structura tertiara
In cazul celulozei, totalitatea problemelor deosebit de complexe adoptata sub denumirea de structura tertiara, structura supramoleculara, structura electrono-microscopica se refera in principal la:
configuratia si dispozitia reciproca a catenelor macromoleculare de celuloza;
caracterul legaturilor dintre catene;
agregarea catenelor de celuloza cu formarea asa-numitelor elemente de structura secundara (u.s.s.);
celula elementara cristalografica;
agregarea elementelor de structura secundara;
forma si dimensiunile acestor agregate supramoleculare;
structura lor interioara si orientarea acestora fata de axa fibrei.
Pentru polimeri, dupa cum si celuloza este un polimer, in stare solida, topita sau in solutii se preconizeaza, de regula, patru tipuri fundamentale de structuri si anume:
structura globulara, in care catenele macromoleculare ale polimerului sint impislite sub forma de ghem;
structura striata, in care catenele macromoleculare ale polimerului se impletesc sub forma striata, aceasta fiind specifica polimerilor inalt elastici;
structura fibrilara, in care catenele macromoleculare ale polimerului sint asociate in pachete, respectiv formatiuni denumite microfibrile sau asociatii ale acestora;
structura partial cristalina, constind din sferolite, agregate sferolite sau monocristale.
Celuloza este un polimer natural cu structura fibrilara, pe cind derivatii celulozici si celuloza regenerata sau mercerizata, prezinta o structura partial cristalina.
3.3.1. Structura cristalina si polimorfizmul celulozei
Unitatile elementare de anhidro-b-D-glucopiranoza din catenele macromoleculare ale celulozei au conformatia spatiala scaun. Mai multe catene de acest fel se asociaza sau se impacheteaza intre ele, prin alipirea planurilor superioare si inferioare imaginare ale panglicilor de celuloza (vezi fig. 3). In acest mod rezulta o formatiune numita pachet de catene macromoleculare (fig. 6). Portiunile hasurate reprezinta regiunile in care practic sint dispuse grupele alcoolice primare -CH2OH, precum si cele secundare libere existente de-a lungul catenelor. Planurile superioare si inferioare imaginare ale celor sapte panglici de celuloza, reprezentate in acest caz in pozitie verticala, sint paralele cu axele acestora.
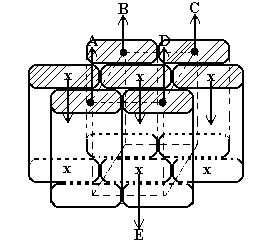
Fig. 6. Reprezentarea schematica a unui pachet alcatuit din sapte catene macromoleculare de celuloza.
Un asemenea mod de impachetare a catenelor macromoleculare de celuloza prezinta urmatoarele particularitati structurale:
Sensul de crestere (de la grupa -OH marginala nereducatoare a primei unitati elementare de structura din stinga panglicii la cea -OH finala reducatoare, aflata in ultima unitate elementara de structura din dreapta panglicii) a catenelor marginale (A,B,C,D) este diferit fata de cel al catenelor centrale (E), sagetile din figura indicind tocmai aceasta alternanta;
Catenele centrale (E) sint deplasate spre stinga sau spre dreapta cu o jumatate de unitate elementara de structura fata de catenele marginale, datorita unor considerente sterice cerute de conformatia tip scaun C1.
Cercetarile röentgenografice au dovedit ca agregatul macromolecular de tipul pachetului de catene constituie de fapt un cristal imaginar foarte mic, orientat paralel cu axa fibrei si se numeste celula elementara cristalina (c.e.c.).
Celula elementara cristalina reprezinta cel mai mic volum posibil al unei retele spatiale cristaline, care pastreaza inca in intregime toate particularitatile geometrice ale structurii in ansamblu.
Modelul acestei celule elementare cristaline imaginare a fost propus si elaborat pentru prima data de catre K. Meyer si H. Mark (1928-29), iar apoi perfectionat de K. Meyer si L. Misch in anul 1937. Acest model este si in prezent acceptat unanim pentru celula elementara cristalina din celuloza nativa, notata conventional prin termenul de celuloza I (fig. 7).
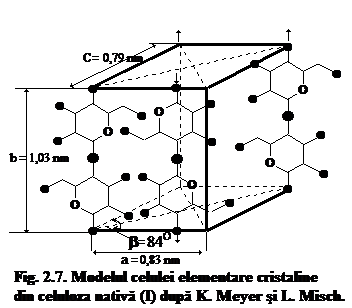
Dimensiunile celulei elementare cristaline au fost evaluate cu ajutorul razelor X si prezinta urmatoarele valori: a = 0,835 nm; b = 1,03 nm; c = 0,79 nm. Unghiul b cuprins intre laturile a si c, care sint perpendiculare pe latura b, are valoarea 84o. La constituirea acestui model, distantele dintre atomii de carbon s-au considerat egale cu 0,154 nm, iar distantele dintre atomii de carbon si oxigen cu 0,135 nm. Planurile unitatilor de anhidro-b-D-glucopiranoza sint dispuse in planul a-b al celulei elementare cristaline, iar axele catenelor macromoleculare de celuloza sint orientate de-a lungul muchiilor b.
Din modelul celulei elementare cristaline (fig. 7) rezulta ca de-a lungul muchiei b se gasesc formatiuni structurale identice cu lungimea de 1,03 nm, care, asa cum s-a mai aratat, concorda perfect cu lungimea unitatii structurale de baza din catena macromoleculara de celuloza, adica a moleculei de celuloza.
Toate celulozele naturale prezinta un singur tip de celula elementara cristalina, care sint proprii celulozei native I, in timp ce diferitele celuloze tehnice modificate, pentru care s-a adoptat notatia celuloza II, cum ar fi hidroceluloza, celuloza mercerizata si celulozele regenerate din solutii, poseda o alta celula elementara cristalina, care este modificata. Simpla tratare, de exemplu, a celulozei native I cu o solutie de NaOH 17,5 %, denumita si mercerizare, urmata de eliminarea alcaliilor cu apa, determina trecerea structurii cristaline intr-o noua forma modificata, care se observa usor si pe difractograma. De aici rezulta si caracterul de polimorfizm al celulozei. In prezent se considera ca röentgenogramele celulozelor tehnice fabricate prin dezincrustarea lemnului, reprezinta imaginea röentgenografica a unui amestec de celuloza I si celuloza II.
Analizind structura celulei elementare cristaline (fig. 6, 7), se poate constata ca in interiorul acestui microcristal imaginar se gasesc cinci catene macromoleculare de celuloza, respectiv patru catene care trec prin muchiile ei si care au sensul de crestere de jos in sus (A,B,C,D), iar la intersectia diagonalelor planului a-c se gaseste cea dea cincia catena macromoleculara cu sensul de crestere opus celorlalte (E). Distantele dintre catenele macromoleculare de celuloza favorizeaza aparitia diferitelor tipuri de legaturi intre atomi sau grupele functionale ale unitatilor elementare de anhidro-b-D-glucopiranoza constituente. Astfel de-a lungul muchiilor b unitatile elementare se mentin legate prin legaturi C1-C4 b-glicozidice. Pe directia muchiilor a se pot forma destul de usor legaturile de hidrogen intermoleculare, deoarece ciclurile de glucoza se gasesc unele fata de altele la o distanta de 0,25 nm. De-a lungul muchiilor C, unde distanta cea mai mica dintre atomi este de cca. 0,31 nm, actioneaza legaturile de tip Van der Waals.
2.3.3.2. Organizarea supramoleculara a celulozei
Dintre primele reprezentari ale structurii supramoleculare a celulozei, se citeaza cea a lui C.Nägeli care, pe baza proprietatilor de umflare si de dubla refractie a celulozei, atribuie asocierea catenelor macromoleculare de celuloza intr-o serie de formatiuni numite micele sau cristalite submicroscopice, implantate intr-o substanta intermicelara cu structura amorfa. De precizat ca prin notiunea de cristalinitate se intelege o orientare cit mai paralel posibila intre catenele macromoleculare de celuloza de-a lungul micelelor. Termenul de substanta intermicelara, cu aspect amorf, se refera in acest caz tot la un ansamblu de catene celulozice, situate insa in exteriorul micelelor si care nu mai sint orientate strict paralele unele fata de altele, ci sint dispuse in mod haotic in spatiul tridimensional.
Conform acestei ipoteze, micelele sint conturate sub forma unor formatiuni paralelipipedice care se dispun in structura fibrelor celulozice analog caramizilor dintr-o zidarie, iar intre micele se afla substanta celulozica amorfa sau "mortarul" intermicelar. Se considera ca lungimea catenelor macromoleculare de celuloza dintr-o micela este limitata la dimensiunile a 30 - 40 unitati elementare de anhidro-b-D-glucopiranoza.
Reprezentarile lui C.Nägeli au fost dezvoltate ulterior de catre H. Mark, K. Meyer si L. Misch, care au aratat ca aceasta organizare supramoleculara ar explica cele doua categorii principale de reactii chimice ale celulozei, respectiv reactiile topochimice si cele micelare. In reactiile topochimice actiunea agentului chimic se limiteaza la suprafata micelelor, distrugind doar substanta amorfa intermicelara, in timp ce in reactiile micelare, reactivul chimic patrunde in profunzimea materialului fibros, cu dislocarea integrala a micelelor si dispersarea acestora in solutie.
Lungimea catenelor macromoleculare de celuloza din micelele imaginate de H. Mark, K. Meyer si L. Misch, este de ordinul a 50 unitati elementare de anhidro-b-D-glucopiranoza, respectiv de cca. 25 nm
In aceeasi perioada, H. Staudinger formuleaza un punct de vedere total diferit. El demonstreaza prin studii viscozimetrice existenta unor lungimi mult mai mari pentru catenele macromoleculare de celuloza din aceste formatiuni primare de organizare supramoleculara. Din acest motiv, el imagineaza structura celulozei ca fiind asemanatoare cu cea a unui polimer in totalitate cristalin, in care catenele macromoleculare de celuloza ar fi intinse si paralele (fig. 8).
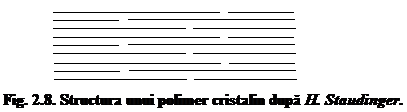
Lungimea catenelor macromoleculare de celuloza dupa H. Staudinger este de ordinul cel putin 800 unitati elementare de anhidro-b-D-glucopiranoza, ajungindu-se astfel la un GP de cel putin 2000 pentru celuloza nativa si de 400 - 800 pentru celulozele regenerate. Lungimea catenelor macromoleculare de celuloza depaseste in acest caz categoric dimensiunile micelelor prezentate anterior, rezultind astfel valori cuprinse 200 - 400 nm 1000 nm.
Plecind de la aceste considerente, ipotezele privind structura tertiara a celulozei au demarat de la bun inceput pe doua planuri distincte si contradictorii. Primul se refera la lungimea catenelor macromoleculare de celuloza, iar cel de al doilea - la modul lor de asociere in structura fibrei de celuloza ca atare. Din acest ultim punct de vedere, se pune intrebarea daca macromoleculele de celuloza se asociaza intre ele in formatiuni supramoleculare distincte de tipul cristalelor, cimentate intre ele de o substanta celulozica necristalina (H.Mark) sau, dimpotriva, intreaga structura fibrilara este continua si omogena, considerata pina la nivel molecular. (modelul H. Staudinger). In ultimele decenii s-au facut numeroase studii experimentale si s-au propus multiple ipoteze, dar pina in prezent nu s-a ajuns la o teorie unitara si definitiva privind structura tertiara a celulozei. S-a reusit totusi sa se grupeze toate aceste ipoteze pentru o expunere mai clara. Astfel Gh. Rozmarin a inclus aproape toate ipotezele in doua grupe mari si anume:
Ipoteze privind structura monofazica a celulozei;
Ipoteze privind structura bifazica a celulozei.
A. Ipotezele structurii monofazice a celulozei se bazeaza in principal pe faptul ca intreaga structura fibrilara a celulozei este continua si omogena. Cele mai importante din acest grup sint: ipoteza structurii total cristaline a celulozei, elaborata de H.Mark, iar mai tirziu de A.Peterlin si P.Ingram; ipoteza structurii paracristaline a celulozei, elaborata de R.Hoseman si prezentata de Gh. Ruscher; ipoteza structurii continue total amorfe a celulozei, elaborata de V.A. Karghin; ipoteza mezomorfizmului celulozei, elaborata de A.I. Kitaigorodski si D. Ja. Tvankin.
B. Ipotezele structurii bifazice a celulozei se bazeaza pe considerarea ca in fibrele de celuloza exista concomitent zone cristaline si zone amorfe. In zonele cristaline catenele macromoleculare sint dispuse ordonat, respectiv paralel intre ele. In zonele amorfe ele sint dispuse in mod dezorganizat, neparalel. Cele mai importante ipoteze din acest grup sint: ipoteza structurii micelare a celulozei are ca punct de plecare modelul lui C. Nägeli. Ea presupune existenta unor formatiuni submicroscopice de tipul micelelor sau a cristalitelor, separate intre ele printr-o substanta celulozica intermicelara de cimentare; ipoteza structurii micelelor franjurate a celulozei elaborata de A.Frey-Wyssling porneste de la faptul ca in spatiul dintre doua micele sint asezate asa-numitele zone franjurate, adica domeniile amorfe, alcatuite din capetele mai multor catene macromoleculare, care apartin uneia sau mai multor micele invecinate; ipoteza structurii microfibrilare a celulozei la elaborarea careia au contribuit mai multi cercetatori; ipoteza structurii fibrilelor franjurate a celulozei elaborata de J.J.Hearle, care incearca sa imbine teoriile structurii micelare a celulozei cu cele microfibrilare;
In anul 1963, in localitatea Baden-Baden din Germania a avut loc un congres consacrat indeosebi problemelor de sinteza si structura supramoleculara a celulozei. Aici s-au formulat oficial o serie de teze, care constituie un punct de vedere sistematizat asupra unor notiuni de terminologie. Ele au fost sintetizate astfel:
Celuloza este un polimer natural cu o structura policristalina, adica cu o structura mai mult sau mai putin ordonata, in care o catena macromoleculara poate participa la alcatuirea mai multor regiuni. Desi tranzitia intre domeniile cristaline si cele amorfe este treptata, structura celulozei este polifazica, deoarece atit in domeniile cristaline, cit si in cele amorfe se pot intilni si spatii goale.
Cristalitul care reprezinta un cristal cu defecte de retea, constituie elementul de baza din asa-numita celuloza cristalina, atunci cind defectele sint minime si, deci, orientarea sau paralelismul catenelor este maxim, si respectiv celuloza paracristalina sau amorfa, atunci cind defectele sint maxime si, deci, orientarea catenelor este minima. Raportul lungimilor laturilor unui astfel de cristalit este estimat la 1:3:20 (3:9:60 nm). Ultima valoare caracterizeaza lungimea cristalitului si este cea mai discutabila.
Raportul dintre marimea regiunilor cristaline si cele amorfe se exprima prin gradul de cristalinitate, care constituie o caracteristica a fiecarui tip de celuloza. Pentru celuloza nativa acesta poate ajunge, de exemplu, la 60 - 70 %.
In ceea ce priveste dispunerea catenelor macromoleculare de celuloza in cristalite, aceasta poate fi de tip: franjurat, cutat sau de tip intermediar. Tipul franjurat a fost demonstrat experimental, iar tipul cutat a fost propus prin analogie cu polimerii sintetici, dar nu s-a demonstrat practic decit partial. Tipul intermediar este doar suficient de probabil.
Cristalitele sint, deci, cele mai mici agregate formate din catena macromoleculara de celuloza, care au putut fi observate cu ajutorul microscopului electronic.
Catenele macromoleculare de celuloza au capacitatea de a se constitui prin asociere intr-o serie de cordoane cristaline sau fibrilare, numite uneori si filamente sau fire micelare, care constau din domenii succesive ordonate sau neordonate (amorfe). Suprafata unui asemenea cordon cristalin este acoperita cu o masa polimera amorfa, dimensiunile acesteia fiind de 6-8 nm in diametru si de 2-5m in lungime. Pina in prezent nu s-a demonstrat inca, daca un asemenea tip de cordon cristalin reprezinta un element nativ de structura supramoleculara sau este rezultatul unor actiuni preparative analitice.
Microfibrila. Agregarea catenelor macromoleculare de celuloza nu se opreste insa la formatiunea de micela sau cristalit, de cordon cristalin sau fibrilar, respectiv de fir micelar sau fibrila elementara, ci prin reunirea mai multor asemenea formatiuni structurale se pot obtine agregate macromoleculare mai complexe denumite microfibrile. Cu toate deosebirile de vederi existente pina in prezent asupra dimensiunilor microfibrilelor, cea mai importanta concluzie care se poate totusi desprinde, este caracterul fibrilar al tuturor fibrelor celulozice, indiferent de originea lor.
Fibrilele. La rindul lor, microfibrilele sint elemente constituente ale altor agregate supramoleculare, mult mai mari si mai complexe, denumite fibrile. Sub actiunea acizilor minerali, fibrila se destrama in fragmente mai scurte cu lungimea de cca. 50 nm, grosimea fiind cuprinsa intre 7,5 si 10 nm. Fractiunile astfel rezultate sint vizibile numai cu microscopul electronic.
Fibrilele de celuloza legate sub forma unor placi alcatuiesc de fapt fibrele de celuloza. Agregarea catenelor macromoleculare de celuloza in cristali elementari, a acestora in cristalite, apoi in microfibrile si asa mai departe pina la fibrile, va duce in mod inevitabil la realizarea unei organizari tridimensionale a fibrei de celuloza care se observa cu ochiul liber.
Golurile sau discontinuitatile pot fi intilnite la toate elementele de agregate macromoleculare mai sus prezentate, constituind un fel de fisuri. Ele formeaza in textura generala diverse canale, care sint dispuse in directie longitudinala si transversala. Latimea fisurilor sau mai curind diametrul acestor goluri dintre cristalite, poate atinge valoarea de 1 nm, iar in zonele amorfe pina la 0,2-0,5 nm. In schimb golurile interfibrilare pot avea dimensiuni cuprinse intre 5 si 13 nm.
In scopuri practice, sint necesar de cunoscut marimile spatiilor libere dintre diferitele formatii supramoleculare, precum si dimensiunile moleculare ale unor substante cu care lemnul intra in contact. Ele au urmatoarele valori:
Spatiul interfibrilar 10 nm
Spatiul intermicelar 1 nm
Spatiul capilar (temporar) din peretii membranei celulare 0,4 nm
Molecula de glucoza 0,5 nm
Molecula de apa 0,3 nm
Molecula unui colorant de natura gudronica 1-2 nm.
Daca se raporteaza aceste valori la lemn, care contine in medie 50% celuloza si densitati cuprinse intre 0,1-1,4 g cm3, se obtine o suprafata specifica interioara de 20-280 m2 cm3. Avind in vedere aceste consideratii teoretice si practice este clar ca in explicarea fenomenelor care se produc la tratarea lemnului cu apa sau diferiti agenti chimici, precum si la tratarea, respectiv uscarea acestuia, trebuie sa se tina seama de valoarea ridicata a acestei suprafete specifice interioare.
In prezent ramin totusi in discutie inca doua probleme si anume: antiparalelismul si cutarea catenelor de celuloza. Antiparalelismul este generat de prezenta a doua catene antiparalele in celuloza nativa. El a fost postulat din motive cristalografice si din unele considerente asupra legaturilor de hidrogen intermoleculare. O serie de cercetari se opun conceptului de antiparalelism in celuloza nativa, deoarece, in acest caz, pentru cresterea fibrilelor de celuloza ar fi necesare doua tipuri de enzime.
Cea de a doua problema o constituie ipoteza cutarii catenelor macromoleculare de celuloza in celuloza nativa. Practic s-au adus dovezi numai asupra existentei cutelor numai in celuloza regenerata si nu in cea nativa. Din aceste motive, in prezent, pentru celuloza se poate considera o structura monofazica, cvasicristalina, cu o periodicitate dimensionala de 3,5 nm, adica cu perioade mici care corespund cristalitului si respectiv de 20 nm, adica cu perioade mari care corespund microfibrilei. Ramine insa de ales intre modelele structurale ale cristalitelor elementare. Masurarile, efectuate prin cercetari electrono-microscopice pentru dimensiunile elementelor structurale constitutive ale unei fibre de celuloza, au dus la o serie de valori, care sint prezentate in tabelul
Tabelul 2
Dimensiunile sectiunii transversale si numarul de catene macromoleculare de celuloza din diferitele elemente structurale ale unei fibre de celuloza.
|
Nr. crt. |
Elementul de structura |
Dimensiunile sectiunii transversale |
Numarul de catene macromoleculare |
|
1 |
Catena macromoleculara de celuloza |
0,83 0,39 nm |
1 |
|
2 |
Cristalitul (firul micelar sau micela) |
5 6 nm |
100 |
|
3 |
Microfibrila |
25 25 nm |
000 |
|
4 |
Fibrila |
0,4 0,4 m |
500.000 |
|
5 |
Fibra de bumbac |
314 m |
750.000.000 |
| Contact |- ia legatura cu noi -| | |
| Adauga document |- pune-ti documente online -| | |
| Termeni & conditii de utilizare |- politica de cookies si de confidentialitate -| | |
| Copyright © |- 2025 - Toate drepturile rezervate -| |
|
|
|||
|
|||
Proiecte pe aceeasi tema | |||
|
| |||
|
|||
|
|
|||