|






 |  |

| Medicina | Medicina veterinara | Muzica | Psihologie | Retete | Sport |
Medicina
|
|
Qdidactic » sanatate & sport » medicina Enzime de oxido-reducere si transportul de electroni - reactii de oxido-reducere |
Enzime de oxido-reducere si transportul de electroni - reactii de oxido-reducere
Principalul izvor motor al activitatilor celulare este transportul de electroni, prin care perechi de electroni derivati de la intermediarii ciclului acidului tricarboxilic si din alte substante se scurg pe lantul respirator spre oxigenul molecular, acceptorul final de electroni din respiratie. In urma procesului de fosforilare oxidativa se elibereaza o mare cantitate de energie libera conservata in mare parte sub forma gruparilor fosfat macroergice ale ATP.
Enzimele de oxido-reducere, si in particular acelea care participa in transportul de electroni, sunt in general mai complexe din punctul de vedere al structurii si mecanismului de actiune si deci mai putin cunoscute decat alte clase de enzime.
1. Reactii de oxido-reducere
Reactiile de oxido-reducere (numite deasemenea reactii redox) sunt acele reactii in care are loc un transfer de electroni de la un donor de electroni (agentul reducator) la un acceptor de electroni (agentul oxidant). In unele reactii de oxido-reducere, transferul unuia sau al mai multor electroni este facut prin intermediul transferului de hidrogen; dehidrogenarea este, prin urmare, echivalenta cu oxidarea. Adeseori se folosesc termenii de echivalenta reducatori sau echivalenti de electroni pentru a se referi la electronii sau atomii de hidrogen participanti in oxido-reducere.
Agentii de oxidare sau de reducere actioneaza ca perechi conjugate redox, fiind formate dintr-un donor de electroni si acceptorul sau de electroni conjugat, asa cum acizii Bronsted si bazele functioneaza ca perechi conjugate acid-baza.
Reactii acid-baza:

Dupa cum diferiti acizi se deosebesc prin tendinta lor de a pierde protoni, tot asa diferiti agenti reducatori difera intre ei prin tendinta de a pierde electroni. Tendinta unui agent reducator de a pierde electroni (sau a unui agent oxidant de a castiga electroni) este data de potentialul standard de oxido-reducere, definit ca forta electromotoare(fem), in volti, data de o semicelula in care atat reducatorul cat si oxidantul se gasesc la o concentratie de 1M, la 250C si pH 7,0 in echilibru cu un electrod care poate accepta reversibil electroni de la reducator, figura 1.
Solutia care contine formele oxidate si reduse ale compusului de examinat este plasata in recipientul din stanga; are loc echilibrarea cu electrodul inert. O punte de sare, care contine o sol. de KCl, asigura legatura cu o celula de referinta de fem cunoscuta (dreapta). Din fem totala masurata si fem cunoscuta a celulei de referinta se poate determina fem a celulei din stanga, care contine amestecul de forma oxidata si redusa a compusului de analizat. Cand concentratia formei reduse este egala cu cea a formei oxidate, potentialul celulei din stanga este potentialul standard sau potentialul de oxido-reducere la echivalenta.
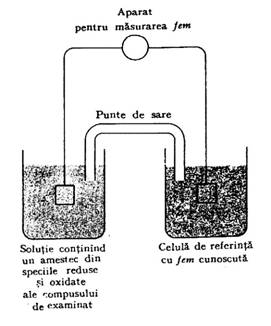
Fig. 1. Masurarea potentialului de oxido-reducere
Prin conventie, ecuatia la electrod este scrisa in sensul:
![]()
unde n este nr. de electroni transferati. De asemenea, tot prin conventie, potentialul standard de oxido-reducere al reactiei la electrodul de hidrogen:
![]()
ste folosit ca potential de referinta. El este considerat 0,0 V cand presiunea H2 gazos este de 1 atm, concentratia H+ de 1M (pH = 0,0) si temperatura de 25 C. Cand aceasta valoare se corecteaza pentru pH 7,0 (aH+s = 1*10-7 M), pH-ul de referinta cerut in toate calculele biochimice, potentialul standard de oxido-reducere al electrodului de hidrogen devine - 0,42 V.
O relatie care exprima legatura dintre potentialul standard de oxido-reducere al unei perechi de oxido-reducere date, potentialul ei observat (masurat), si raportul concentratiilor speciilor acceptoare de electroni si donoare de electroni este ecuatia Nernst:
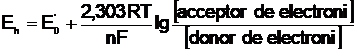
in care:
E0 = potentialul redox standard (pH = 7,0, T =25 C sau 298 K, toate concentratiile componentilor fiind de 1,0M);
Eh = potentialul de electrod observat (masurat);
R = constanta gazelor (8,31 Jgrad-1mol-1);
T = temperatura, a°Ks
n = nr. de electroni care sunt transferati;
F = 23062 cal*V = 96406 J*V-1 - nr. lui Faraday
La 25 C (298K) termenul 2,303 RT/nF are valoarea 0,059 cand n=1 si 0,03 cand n=2. Deoarece se obisnuieste sa se calculeze echilibrele perechilor de oxido-reducere biologice in termeni ai transferului de 2 electroni, ecuatia Nernst se simplifica la:
 2)
2)
Ecuatia Nernst exprima matematic alura unei curbe de titrare a unui donor de electroni dat cu un agent de oxidare puternic, figura 2.
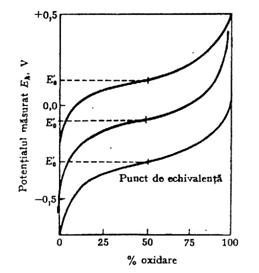
Fig. 2. Curbele de titrare de oxido-reducere a trei perechi redox conjugate avand diferite potentiale standard, sau de echivalenta
Pe masura ce titrarea progreseaza, o fractiune crescanda a donorului de electroni trece in farma sa oxidata avand ca rezultat cresterea raportului aacceptor de electronis / adonor de electronis pana la atingerea punctului de echivalenta, unde aacceptorul de electronis = adonorul de electronis. La acest punct, termenul 0,03 lg aacceptor de electroni s / a donor de electroni s este zero si ecuatia Nernst se reduce la:
|
![]() (3)
(3)
Prin urmare, potentialul standard redox este forta electromotoare in volti a unei pile la exact punctul de echivalenta a unei curbe de titrare a unui reducator dat, la pH 7,0, 25 C si 1,0 atm. dupa cum pK'-ul unui acid este egal cu pH-ul la punctul de echivalenta pe curba de titrare a unui acid cu o baza. Potentialul standard redox, E0, este adesea numit potential de echivalenta. Spre deosebire de titrarile acido-bazice, care sunt reactii ionice rapide ce nu necesita catalizatori, reactiile de oxido-reducere ale compusilor organici sunt mai degraba incete si necesita prezenta unui catalizator sau a unei enzime pentru atingerea rapida a echilibrului.
Potentialele standard de oxido-reducere, ale diferitelor sisteme biologice de oxido-reducere permit precizarea directiei in care electronii au tendinta sa treaca de la o pereche redox la alta in conditii standard.
Concentratiile la echilibru ale celor patru componenti dintr-o reactie de oxido-reducere pot fi calculate din potentialele standard ale sistemelor care interactioneaza si din concentratiile totale ale tuturor componentelor.
2. Clase de enzime transportoare de electroni
Patru tipuri de enzime de oxido-reducere sau proteine care transfera electroni participa in fluxul principal al transportului de electroni de la substrate organice la oxigenul reducator. Ele sunt:
dehidrogenazele legate de nucleul piridinic (piridin-dependente) care necesita NAD sau NADP drept coenzime;
dehidrogenazele legate de nucleul flavinic (flavin-dependente), care contin flavinadenin dinucleotidul (FAD) sau flavin mononucleotidul (FMN) ca grupare prostetica;
proteinele cu sulf;
citocromii, care contin inelul porfirinic cu Fe ca grupare prostetica.
In plus fata de aceste proteine, in transportul de electroni functioneaza si coenzima solubila in lipide numita ubichinona sau coenzima Q.
2.1. Dehidrogenaze legate de piridina (piridin-dependente)
Deoarece membrii acestei clase de dehidrogenaze necesita NAD sau NADP drept coenzime, acestea continand un derivat al piridinei, nicotinamida, ei sunt denumiti generic dehidrogenaze piridin-dependente. Se cunosc circa 200 sau mai multe astfel de enzime care functioneaza in diferite cai metabolice. Aceste enzime catalizeaza reactiile generale:
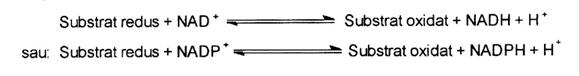
In functie de tipul de piridin nucleotid preferat.
Nicotinamidadenin dinucleotidul (NAD+) si nicotinamidaadenin dinucleotidfosfat (NADP+) sunt coenzime implicate intr-un numar mare de procese redox. Aceste coenzime sunt usor disociabile de apoenzimele lor, avand constante de disociere de ordinul 10-4 - 10-6 M. In general, formele reduse sunt legate mai puternic decat formele oxidate.
NAD este constituit dintr-un rest de adenina, unul de nicotinamida, doua resturi de acid fosforic si doua resturi de riboza legate prin legaturi -glicozidice la atomii de azot a celor doi heterocicli. NADP poseda in plus un al treilea rest fosforic legat la riboza printr-o legatura esterica.
In celule animale, NAD se gaseste in cantitate mult mai mare decat NADP. In ficat, cca. 60% din NAD total se afla in mitocondrie, iar restul se gaseste in citoplasma.
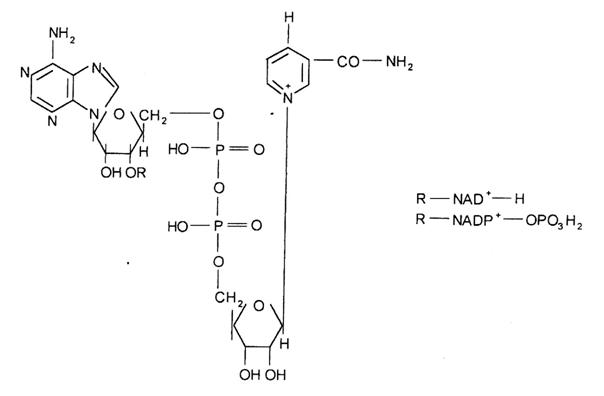
Continutul de NADP al celulelor este proportional cu activitatea lor biosintetica. Dehidrogenazele NAD-dependente au rolul principal in respiratie, pe cand NADP-dependente servesc, in primul rand, la transferul electronilor de la intermediari ai catabolismului la intermediari ai biosintezei. NAD+ si NADP+ liberi se gasesc in solutie in doua conformatii, si anume forma deschisa sau extinsa si forma inchisa sau stransa, in care inelele nicotinamidei si adeninei sunt paralele unul cu altul. Forma stransa predomina in solutie, dar, cand NAD+ se leaga de unele dehidrogenaze, trece in conformatia deschisa. Cele mai multe dehidrogenaze piridin-dependente sunt specifice fie pentru NAD, fie pentru NADP, dar cateva, de ex. glutamat dehidrogenaza, pot reactiona cu ambele coenzime.
Multe dehidrogenaze piridin-dependente contin strans legati ioni metalici divalenti; de exemplu alcool dehidrogenaza contine Zn2+.
Mecanismul de actiune al unor piridinnucleozid enzime-alcool dehidrogenaza
Enzima din drojdie de bere are o masa moleculara de 151000 daltoni, contine patru atomi de zinc per molecula si leaga patru molecule de NAD+, in timp ce enzima omoloaga din tesutul hepatic are o masa de 84000 daltoni, leaga numai doi atomi de zinc si doua molecule de NAD+. Indepartarea ionului metalic prin tratament cu diversi agenti de complexare provoaca o inactivare insotita si de o disociere a proteinei in subunitati, ceea ce indica rolul dublu al metalului-structural si functional. Atomul metalic este implicat in legarea NAD+ si substratului la centrul activ al enzimei si in ansamblarea celor patru subunitati proteice in forma tetramera, nativa. Dislocuirea NAD+ cu p-mercuribenzoat si protejarea enzimei cu NAD+ fata de actiunea iodacetamidei indica prezenta unei grupari sulfhidril in vecinatatea coenzimei legate.
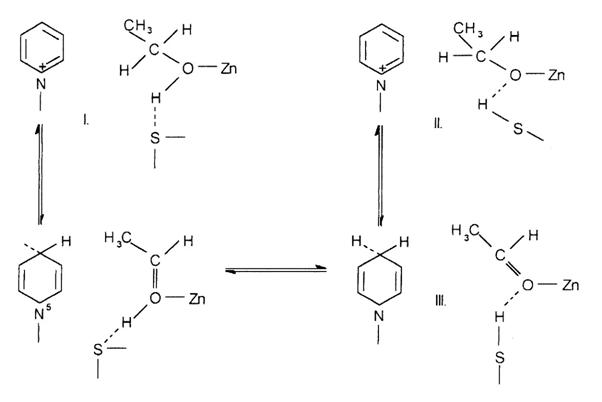
Deoarece in medii cu constante dielectrice mici, 1-metil-nicotinamida si anionii formeaza perechi de ioni stabili se asteapta ca legarea NAD+ la alcool dehidrogenaza sa aiba ca rezultat formarea unei perechi de ioni implicand atomul de azot incarcat pozitiv de la NAD+ si atomul de sulf incarcat pozitiv al unei grupari SH vecine. Daca se presupune ca etanolul este legat coordinativ la Zn2+ are unul din atomii de hidrogen orientat pentru transfer stereospecific fie spre atomul C4 din ciclul piridinic al coenzimei si cu gruparea metil libera de a se roti, conform structurii I in care gruparea HO a etanolului este legata prin intermediul hidrogenului la o grupare SH a apoenzimei. Transportul de hidrogen facilitat de-a lungul acestei legaturi de hidrogen aduce structura I intr-un echilibru rapid cu structura II. Aceasta din urma este termodinamic instabila datorita sarcinii ei net pozitive, are tendinta de a scadea, printr-un efect inductiv, densitatea de electroni la atomul C4, dupa cum indica sagetile. intre timp, atomul de oxigen activat al ionului alcoxid tinde a forma o legatura in plus cu propriul lui atom de carbon si deci de a slabi legatura vecina C - H. Aceste doua efecte coopereaza in transferul ionului hidrura care transforma structura II si III. Invers, in reactia inversa de hidrogenare, III se poate transforma in II prin transferul ionului hidrura si apoi in I prin transferul facilitat de protoni. Deoarece transferurile de hidrura si de proton sunt legate, intre ele exista o relatie reciproca si nici unul nu-l poate precede pe celalalt.
2.2. Dehidrogenaze legate de flavina (flavin-dependente)
Flavinenzimele sunt proteine care in general contin puternic legate in structura lor gruparile prostetice: flavin mononucleotidul (FMN) si flavinadenin dinucleotidul (FAD), care prezinta un nucleu 7,8-dimetil izoaloxazinic, sediul proceselor redox, o catena ribitol purtand o functiune ester fosforic sau o grupare adenozin monofosfat:
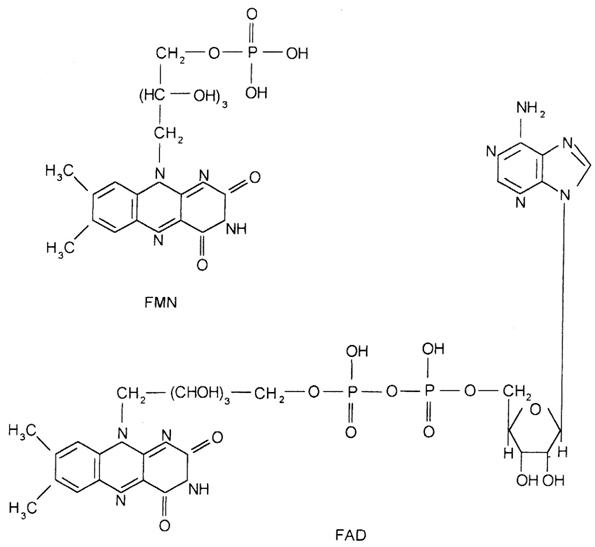
Partea activa a FMN sau FAD, care participa la reactia de oxido-reducere, este ciclul izoaloxazinei din partea riboflavinei, care este redus reversibil:
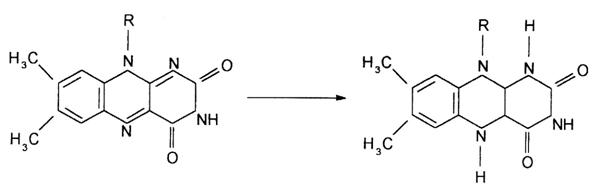
In acesta reactie are loc transferul direct al unei perechi de atomi de hidrogen de la substrat pentru a genera formele reduse ale FMN si FAD, notate cu FMNH2 si FADH2.

Cele mai importante dehidrogenaze flavin-dependente in curentul principal al respiratiei si transportului de electroni, toate localizate in mitocondrie, sunt:
NADH dehidrogenaza, care coiitine FMN si catalizeaza transferul electronilor de la NADH la urmatorul membru al lantului transportor de electroni;
succinat dehidrogenaza, activa in ciclul acizilor tricarboxilici;
dihidrolipoil dehidrogenaza, un component al sistemelor piruvat si a-cetoglutarat dehidrogenazelor;
acil-CoA dehidrogenaza, care catalizeaza prima etapa de dehidrogenari din oxidarea acizilor grasi.
Dehidrogenazele flavin-dependente difera semnificativ de acelea piridin-dependente prin aceea ca flavin nucleotidul este foarte strans legat de molecula enzimei si astfel functioneaza mai degraba ca grupare prostetica decat ca o coenzima; flavin nucleotidul nu paraseste enzima in timpul sau dupa efectuarea ciclului catalitic. in cele mai multe flavin dehidrogenaze, incluzand NADH dehidrogenaza, flavin nucleotidul este legat necovalent; constantele de disociere sunt in domeniul 10-8-10-11M. Astfel de flavoenzime pot uneori fi disociate prin expunere la tarii ionice ridicate sau pH mic.
Enzimele de oxido-reducere flavin-dependente pot fi plasate in doua clase, dehidrogenaze si oxidaze, in functie de capacitatea lor de a reactiona cu acceptorii de electroni.
Diferitele flavin dehidrogenaze in forma complet oxidata sunt galbene, rosii, brune sau verzi; ele au de obicei maxime de absorbtie in zona 370 si 450nm. Cand sunt complet reduse, enzimatic sau chimic, ele isi pierd culoarea si maximul de absorbtie la 450 nm. Schimbari caracteristice ale fluorescentei au loc, de asemenea, in timpul ciclului de oxido-reducere al unor flavin enzime.
Desi unele flavoproteine oscileaza intre forma complet oxidata si cea complet redusa printr-un transfer simultan de doi electroni, alte flavin enzime par sa transfere numai cate un electron odata in timpul ciclului catalitic normal si astfel pot scinda perechile de electroni. Transferul unui singur atom de hidrogen sau electroni pe o molecula de FAD sau FMN (sau pierderea unui hidrogen sau a unui electron de la o molecula de FADH2 sau FMNH2) conduce la formarea unei forme semireduse, sau semichinonice, a flavin nucleotidului. Aceste forme, care sunt radicali liberi, pot fi detectate fie prin spectrele lor de absorbtie caracteristice, fie prin spectroscopia de rezonanta electronica de spin, care semnaleaza prezenta radicalilor liberi, de ex. molecule cu spin electronic nepereche, si anume prin urmarirea comportarii lor caracteristice intr-un camp magnetic.
Unele flavoproteine contin, pe langa flavin nucleotid, si metale, in general fier si molibden; aceste metale sunt esentiale pentru exprimarea activitatii catalitice.
Mecanismul de actiune al unor flavin enzime - succinat dehidrogenaza
In aceasta flavin enzima, flavin nucleotidul este legat covalent printr-o legatura intre ciclul izoaloxazinic al FAD si un rest de aminoacid al proteinei.
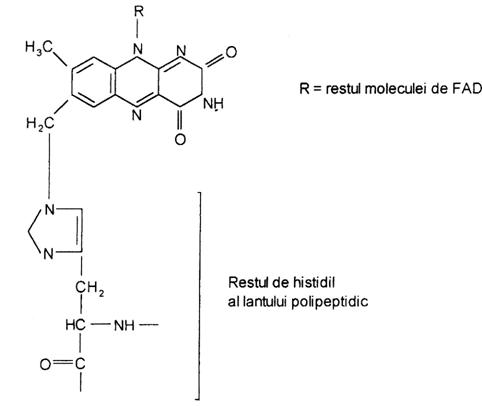
Succinat dehidrogenaza catalizeaza oxidarea succinatului la fumarat si contine legat covalent in structura proteica FAD care actioneaza ca acceptor de hidrogen, in reactia:
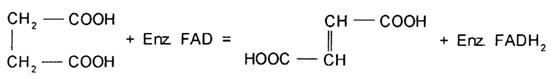
Enzima redusa poate ceda electroni unor acceptori de electroni artificiali (coloranti reductibili), acceptorul fiziologic normal nefiind cunoscut cu certitudine. Enzima este membru al ciclului Krebs la celulele aerobe, ca cele ale vertebratelor, drojdiilor aerobe sau plantelor superioare, poseda o mare afinitate pentru succinat si o mica afinitate pentru fumarat, astfel incat acumularea ultimului substrat nu inhiba reactia. Succinat dehidrogenaza de la toate celulele aerobe examinate, cand vine in contact cu substratul, sufera o transformare conformationala, rapida si reversibila de la o forma cu slaba activitate catalitica la alta extrem de reactiva. Acest fenomen de 'activare' are, probabil, un rol reglator in oxidarile mitocondriale. Prin intermediul atomilor de Fe neheminic si al lipidelor asociate, succinat dehidrogenaza realizeaza un contact functional deosebit de eficient cu ubichinona si prin intermediul acesteia cu sistemul citocromic. Ea nu este autooxidabila nici la presiuni mari ale O2. Deoarece este puternic legata de membrana mitocondriala, succinat dehidrogenaza este dificil de extras si purificat si nu exista certitudinea obtinerii unui preparat cu conformatie si structura chimica identice celei native. Astfel, purificata din suspensii mitocondriale izolate din miocard bovin, enzima pare a avea o organizare subunitara complexa cu o masa moleculara de aprox. 175000 daltoni, cu un mol de FAD legat covalent, cu patru atomi de fier si patru atomi de sulf. Exista opinia ca in timpul catalizei enzimatice fierul sufera un proces de oxidare. Atomii de fier si sulf fac parte din asa-numita categorie a feroproteinelor neheminice. Acesta feroproteina neheminica pare a avea o masa de 26000 daltoni si cate doi atomi de Fe si sulf 'labil' per molecula.
Sterochimia indepartarii atomilor de hidrogen din structura succinatului a fost studiata prin experimente cu deuteriu si cu clorosuccinat:
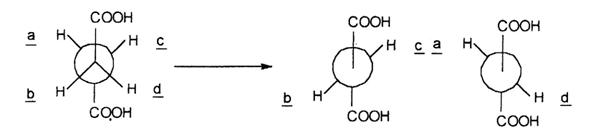
Succinat dehidrogenaza catalizeaza cu viteze de reactie egale trans-dehidrogenaza, indepartarea atomilor de hidrogen din pozitii trans (b si c) - (a si d). Enzima manifesta unele proprietati care amintesc de enzimele alosterice fiind inhibata competitiv de concentratii mici de oxaloacetat sl activata de fumarat, succinat si fosfat.
Enzime cu fier-sulf
Proteinele cu fier-sulf contin fier si sulf liberabil cu acizi, in cantitati echimoleculare. Prima proteina cu fier-sulf descoperita, ferodoxina, a fost identificata intr-o bacterie anaeroba, Clostridium pasteurianum, capabila de a fixa azotul atmosferic. Proteine asemanatoare au fost descoperite apoi si in plante superioare, unde ele se gasesc in cloroplaste si participa la transportul de electroni al fotosintezei. Proteinele cu fier-sulf au fost, de asemenea, gasite in alte microorganisme si in tesuturi animale, in mod special in mitocondrii. Mai mult, grupari Fe - S se gasesc in unele flavoproteine, unde se numesc centri de fier-sulf.
Masa moleculara a proteinelor cu fier-sulf este in domeniul 6000-100000 sau mai mare. Aceste proteine par sa functioneze ca transportori de electroni suferind tranzitii reversibile Fe(II) - Fe(III). Formele Fe(III) sunt rosii sau verzi; prin reducere are loc o decolorare partiala. Formele reduse Fe (II) ale celor mai multe proteine cu fier-sulf arata absorbtia rezonantei electronice de spin la temeraturi joase (4-100 K) cu o valoare electronica g caracteristica intre 1,90 si 2,90 gauss, care arata prezenta electronilor nepereche la forma Fe(II).
In lantul mitocondrial de la NADH la oxigen par sa existe cel putin sapte centre fier-sulf diferite, detectate prin metoda rezonantei electronice de spin. Patru sunt localizate in complexul NADH dehidrogenazei, doua sunt asociate citocromului b si unul citocromului c1. Desi acesti centri sunt fara indoiala foarte importanti in transportul de electroni, functia lor precisa nu este inca cunoscuta.
Citocromii
Citocromii sunt proteine transportoare de electroni care contin grupari porfirinice cu fier; ei sunt prezenti numai in celulele aerobe. Unii sunt localizati in membrana mitocondriala interna, unde actioneaza secvential pentru a transporta electronii proveniti de la diferite sisteme ale dehidrogenazelor pana la oxigenul molecular.
Alti citocromi se afla in reticulul endoplasmatic, unde joaca un rol important in reactiii de hidroxilare specializate. Toti citocromii sufera tranzitii reversibile de valenta Fe(II) - Fe(III) in timpul ciclului lor catalitic. Formele reduse nu pot fi oxidate de oxigenul molecular, cu exceptia citocromului terminal al respiratiei mitocondriale, si anume citocromul a3 sau citocrom c oxidaza, care contine, de asemenea, cupru strans legat. in mitocondriile animalelor superioare, unde lantul respirator a fost cel mai intens studiat, cel putin cinci citocromi diferiti au fost identificati in membrana interna: citocromii b, c1, c, a si a3.
Citocromii au fost descoperiti prima oara, in 1886, de C. MacMunn si denumiti histohematine, dar semnificatia lor in oxidarile biologice nu a devenit clara pana in 1925, cand ei au fost redescoperiti de D. Keilin in Anglia.
Citocromii, cu o singura exceptie, sunt foarte strans legati de membrana mitocondriala si deci sunt dificil de obtinut in stare solubila si de purificat pana la omogenitate. Citocromul c insa se extrage usor din mitocondrii, cu ajutorul solutiilor concentrate de saruri. Citocromul c este singura proteina cu hem in care hemul este legat de proteina printr-o legatura covalenta. In hemoglobina si mioglobina, ca si in citocromii b si a, inelul porfirinic este legat necovalent si poate fi indepartat prin extractie din proteina acidulata, cu piridina sau cu alti solventi.
Citocromii a si a3se gasesc combinati in aceesl molecula oligomerica mare; cele doua grupari hem sunt identice din punct de vedere chimic, insa difera in ceea ce priveste reactivitatea fata de unii liganzi. Acest complex este cunoscut sub denumirea de citocrom aa3. El are o masa moleculara de cea. 200000 si contine un numar de subunitati de diferite marimi moleculare. Enzima contine doua molecule de hem. A si doi atomi de cupru. Electronii sunt primiti de la citocromul c de catre hemul a si sunt apoi transferati hemului a3. Cei doi atomi de cupru ai citocrom c oxidazei, care dau semnale caracteristice de rezonanta electronica de spin sl sufera trazitii Cu(II) - Cu(I) in timpul transportului de electroni, se pare ca participa la transferul electronilor de la hemul a3 la oxigen. Ionul CN- inhiba reoxidarea citocromului a3 de catre oxigen; hidrogenul sulfurat are o actiune asemanatoare. Citocromul a3, ca sl hemoglobina, se combina cu CO, probabil la locul ocupat in mod normal de oxigen. Inhibitia citocrom oxidazei de catre CO este inversata prin iluminare cu lumina vizibila.
2. Ubichinona
Ubichinona sau coenzima Q este o coenzima liposolubila care paricipa, de asemenea, la transportul electronilor de la substrate organice la oxigen, in lantul respirator al mitocondriei. Aceasta coenzima, care este o chinona reductibila in mod reversibil, are o catena laterala lunga, izoprenoida si a fost descoperita pentru prima oara de R.A. Morton, care a numit-o ubichinona datorita raspandirii ei ubicuitare in animale, plante si microorganisme. Mai tarziu, F.L. Crane si colab. au gasit in mitocondrie o chinona liposolubila, pe care au postulat-o ca functionand in transportul de electroni si pe care au denumit-o coenzima Q. Apoi s-a descoperit ca aceasta este identica cu ubichinona.
De fapt se cunosc mai multe ubichinone care difera numai prin lungimea catenei izoprenoide, care are 6 unitati de izopren in unele microorganisme si 10 in mitocondriile tesuturilor animale. In tesuturile vegetale, corespondentele ubichinonelor sunt plastochinonele care indeplinesc functiuni similare in transportul de electroni ai fotosintezei.
| Contact |- ia legatura cu noi -| | |
| Adauga document |- pune-ti documente online -| | |
| Termeni & conditii de utilizare |- politica de cookies si de confidentialitate -| | |
| Copyright © |- 2025 - Toate drepturile rezervate -| |
|
|
|||
|
|||
|
|||
Referate pe aceeasi tema | |||
|
| |||
|
|||
|
|
|||