|






 |  |

| Biologie | Botanica | Chimie | Didactica | Fizica | Geografie |
| Gradinita | Literatura | Matematica |
Biologie
|
|
Qdidactic » didactica & scoala » biologie Ciclul celular |
Ciclul celular
CICLUL CELULAR
Intervalul de timp cuprins intre momentul din care s-a incheiat diviziunea celulara, concretizata cu aparitia unei noi celule pana la terminarea diviziunii proprii, ale celulei nou aparute, constituie ciclul de viata al unei celule sau ciclul celular (grec. kyklos - cerc). Aceasta duplicare si divizare repetata reprezinta mecanismul principal prin care se reproduc toate celulele (Fig. 46).
La organismele unicelulare (procariote), prin fiecare diviziune celulara iau nastere organisme complet noi, in contrast cu organismele pluricelulare, pentru care sunt necesare diviziuni succesive pentru a "produce" un nou organism provenit dintr-o singura celula (celula ou sau zigotul).
Ciclul celular cuprinde doua mari faze:
I. Interfaza sau interkineza, care cuprinde trei faze:
- faza G1 (faza presintetica);
- faza S (de sinteza);
- faza G2 (postsintetica);
II. Diviziunea celulara, care la randul ei, cuprinde:
- mitoza (diviziunea nucleara);
- citokineza (separarea citoplasmei).
Mitoza - 2 ore celula fiica II r

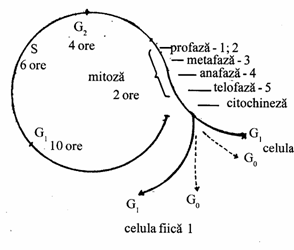
Fig. 46. Fazele ciclului celular (Mariana Sincai, 2000)
G1 sau faza presintetica cuprinde perioada de timp de la sfarsitul mitozei si pana la initierea sintezei de ADN. Pe parcursul acestei faze, celula este metabolic activa, creste continuu si in care are loc o sinteza intensa de ARNm, proteine, enzime, si de molecule macroergice. La o serie de celule, cum sunt celulele tumorale, aceasta faza poate lipsi.
S este faza de sinteza a ADN, cand are loc fenomenul de replicatie a materialului genetic (dublarea cantitatii de ADN);
G2 este faza postsintetica, cand au loc o serie de fenomene pregatitoare ale diviziunii propriu-zise;
M constituie faza de diviziune, in urma careia rezulta cele doua celule fiice in care materialul genetic se va distribui in mod egal, dupa care celulele trec in faza G1 reluandu-se ciclul celular.
Pe toata durata ciclului celular, cantitatea de material genetic nu este reprezentata constant, astfel: in faza G1 - cantitatea de ADN variaza, in faza S - se dubleaza, in faza G2 - ramane constanta, pentru ca in faza M sa se reduca la jumatate. Cu alte cuvinte, celulele se pot distinge in functie de faza ciclului celular in care se afla si pe baza continutului de ADN. De exemplu, celulele somatice in faza G1 sunt diploide - 2n (contin doua molecule de ADN, una de origine materna si una de origine paterna, avand gene pentru aceleasi caractere). Pe parcursul fazei S, in urma replicatiei, creste continutul de ADN al celulei si din diploida devine tetraploida - 4n, adica va contine 4 molecule de ADN, organizate in doi cromozomi. In faza G2, continutul de ADN se mentine - 4n, pentru ca in faza M, dupa citokineza, sa scada, reducandu-se la jumatate - 2n.
Durata ciclului celular variaza considerabil in functie de specie, de tipul si functiile celulei, precum si de conditiile mediului ambiant. Aceasta variatie se remarca numai in ceea ce priveste durata perioadei G1, in care unele celule pot ramane chiar ani de zile. Dupa depasirea fazei G1, perioada de timp necesara pentru parcurgerea fazei S si G2 este relativ constanta pentru toate tipurile celulare. Diviziunea celulara propriu-zisa (faza M) se realizeaza in 1 - 8 ore.
|
Astfel, pentru o celula de tip uman, cu capacitate de proliferare rapida, timpul total al ciclului celular este 24 ore, din care faza G1 se parcurge in aproximativ 11 ore, faza S - 8 ore, faza G2 - 4 ore si faza M - 1 ora.
Exista si alte tipuri de celule care pot parcurge acest ciclu mult mai rapid. Spre exemplu, pentru drojdii (eucariote unicelulare), ciclul celular se deruleaza, cu toate cele patru faze, in aproximativ 90 minute. Pentru celulele embrionare timpurii, ciclul celular este si mai scurt, de 30 minute sau chiar mai putin, si incepe imediat dupa fertilizarea oului sau zigotului. In acest caz, totusi, nu are loc o crestere celulara, prin urmare nu exista fazele G1 si G2, iar replicarea moleculelor de ADN se produce foarte rapid. Practic, ciclul celular al celulelor embrionare timpurii consta intr-o alternanta de faze scurte de sinteza si mitoza.
Reglarea ciclului celular si punctele de restrictie (control). Parcurgerea ciclului celular de catre celule este reglata de o varietate de molecule de semnalizare intracelulare si extracelulare, care monitorizeaza si coordoneaza toate procesele manifestate in acest interval de timp. Un exemplu de reglare a ciclului celular prin semnale extracelulare este indus de catre efectul factorilor de crestere asupra proliferarii celulelor animale. In plus, diferite procese celulare cum sunt: cresterea celulara, replicatia moleculelor de ADN si mitoza, trebuiesc coordonate pe tot parcursul ciclului celular. Aceasta coordonare celulara se realizeaza trecand printr-o serie de puncte de restrictie sau de control, care regleaza parcurgerea fazelor ciclului celular.
Punctele de control sau de restrictie regleaza parcurgerea ciclului celular ca raspuns la dimensiunea celulei sau la moleculele de semnalizare extracelulare, cum sunt nutrientii si factorii de crestere. In plus, evenimentele care au loc in diferitele faze ale ciclului celular trebuiesc coordonate unele in raport cu celelalte pentru ca sa existe o succesiune ordonata si adecvata. De exemplu, este deosebit de important ca celulele sa nu inceapa procesul de mitoza pana nu are loc replicatia completa a genomului. Alternativa acestui fapt ar conduce la desfasurarea unei diviziuni defectuoase, in care celulele fiice nu reusesc sa mosteneasca copiile complete de material genetic.
La majoritatea celulelor, aceasta coordonare intre diferitele faze ale ciclului celular este dependenta de un sistem de puncte de restrictie si de control feedback, care previne intrarea celulei in urmatoarea faza pana cand evenimentele fazei precedente nu au fost incheiate.
Diferitele puncte de restrictie si control functioneaza in scopul de a garanta faptul ca anumiti cromozomi incompleti sau lezati nu se vor replica si, in acest fel, nu vor fi mosteniti de catre celulele fiice.
Moleculele de semnalizare extracelulare pot controla proliferarea celulara, regland parcurgerea ciclului celular atat din faza G1 in faza S, cat si din faza G2 in faza M.
Pentru majoritatea celulelor animale si vegetale, proliferarea este reglata inca din faza G1 a ciclului celular.
Specific pentru celulele animale este punctul decizional din faza G1 tarzie, denumit si punct de restrictie (punct R). Existenta punctului R apare ca o adaptare a celulelor in scopul de a supravietui atunci cand conditiile de mediu sunt nefavorabile si/sau sinteza proteinelor este redusa, dar suficienta pentru supravietuire.
Parcurgerea ciclului celular de catre celulele animale este reglata de factorii de crestere extracelulari, care semnalizeaza proliferarea celulara si, intr-o mai mica masura, de disponibilitatea nutrientilor. In prezenta factorilor de crestere adecvati, celula poate trece de punctul de restrictie si intra in faza S a ciclului celular. Practic, odata depasit punctul R, celula se obliga sa incheie fazele ciclului celular, chiar si in absenta unei stimulari ulterioare a factorilor de crestere. Pe de alta parte, parcurgerea ciclului celular este oprita in punctul de restrictie, daca factorii de crestere adecvati nu sunt disponibili in faza G1. Astfel de celule "deconectate" intra intr-o faza statica a ciclului celular, denumita faza G0, in care pot ramane pentru perioade lungi de timp. Cu toate ca procesul de creste este oprit, sinteza proteica este redusa, celulele din faza G0 sunt metabolic active. La animale, multe tipuri de celule raman in faza G0, daca nu primesc semnale de proliferare, prin intermediul factorilor de crestere adecvati sau alte molecule de semnalizare extracelulare. De exemplu, din necesitatea repararii alterarilor induse de rani, fibroblastele din derma, oprite in faza G0, sunt stimulate pentru a intra in diviziune. Proliferarea acestor fibroblaste este declansata de catre factorii plachetari de crestere, care sunt eliberati de trombocitele sangvine in timpul procesului de coagulare si care induc multiplicarea celulelor din imediata vecinatate a tesutului lezionat.
Cauza care determina trecerea celulelor dincolo de punctul R nu este suficient cunoscuta.
Una din teoriile acceptate afirma ca aceasta parcurgere a fazei G1 si trecerea punctului R are loc in urma acumularii in celule, intr-o anumita concentratie, a unei proteine denumite proteina unstable sau U, declansandu-se astfel si sinteza moleculelor de ADN. Aceasta teorie explica in parte de ce perioada G1 se prelungeste atunci cand sinteza proteinelor este inhibata. Astfel, cand proteina U se sintetizeaza in cantitate mica, celula nu intra in diviziune dar ramane viabila pe perioade lungi, chiar in conditii de aport extrem de redus de substante nutritive.
O alta teorie sustine existenta in celule a unei proteine care inhiba mitoza numita proteina inhibitorie M, care, intr-o anumita concentratie, impiedica celulele sa treaca de punctul R. Aceasta substanta inhibitorie M este sintetizata constant de catre celule si prezinta un turnover rapid. In momentul cand volumul citoplasmatic creste, concentratia proteinei inhibitoare M se va diminua in volumul citoplasmatic crescut si efectul ei inhibitor este anulat, celulele putand sa depaseasca punctul R si sa intre in diviziune. Investigatiile recente au relevat ca proteina 68 kD, sintetizata in G1 ar indeplini in celule rolul proteinei inhibitoare M.
Desi, pentru majoritatea celulelor, proliferarea este reglata in principal in faza G1, exista anumite cicluri celulare care sunt controlate mai ales in faza G2. In acest sens, pentru citologia animala, exemplul elocvent de ciclu celular controlat in faza G2 este oferit de celulele gonozomale (sexuale). Astfel de celule pot ramane blocate in faza G2 pentru perioade foarte lungi de timp (ex.: cateva decenii la om), pana in momentul in care are loc stimularea hormonala, declansand parcurgerea fazei M.
Puncte de control din faza G2 sesizeaza moleculele de ADN nereplicate si genereaza semnale care franeaza ciclul celular. Din acest motiv, in urma activitatii punctelor de control G2, se previne initierea fazei M inainte ca faza S sa fie completa, celulele ramanand in aceasta faza pana cand genomul va fi replicat complet. Numai atunci cand genomul este complet replicat are loc parcurgerea fazei G2, permitand initierea mitozei si distribuirea cromozomilor in celulele fiice.
De asemenea, parcurgerea ciclului celular este oprita in punctele de control G2 in urma lezarii moleculelor de ADN, ca rezultat al iradierii. In majoritatea cazurilor, stationarea in faza G2 asigura timpul necesar pentru repararea leziunilor produse in structura ADN-ului[1], inainte ca acestea sa fie transmise celulelor fiice.
Un alt punct de control al ciclului celular, care mentine integritatea genomului, apare la sfarsitul mitozei. Acest punct de control monitorizeaza alinierea cromozomilor pe fusul de diviziune, asigurand astfel distributia cu acuratete a fiecarui set complet de cromozomi pentru celulele fiice. De exemplu, esecul alinierii exacte a unuia sau a mai multor cromozomi pe fusul de diviziune determina oprirea mitozei in metafaza, inaintea separarii cromozomilor in cei doi nuclei ai celulelor fiice. Ca rezultat final al actiunii acestui punct de control, cromozomii nu se separa pana in momentul in care perechea lor complementara nu este organizata pentru distributia in fiecare celula fiica.
Cuplarea fazei S cu faza M. Punctele de control din faza G2 previn initierea mitozei inainte de a se finaliza faza S, prin aceasta garantand faptul ca moleculele de ADN incomplet replicate nu vor fi repartizate in celulele fiice. La fel de importanta este, de asemenea, garantarea faptului ca genomul sa se replice o singura data per ciclu celular. Cu alte cuvinte, odata ce ADN-ul s-a replicat, este necesar sa existe mecanisme de control care sa previna initierea unei noi faze S, inaintea mitozei. Aceste puncte de control previn celulele din faza G2 sa reintre in faza S si blocheaza initierea unui alt ciclu de replicare a moleculelor de ADN pana dupa mitoza, punct la care celulele intra in faza G1 a urmatorului ciclu celular.
In urma experimentelor efectuate pe fuziuni celulare s-a demonstrat ca atunci cand celulele din faza G1 fuzioneaza cu celule din faza S, nucleul G1 incepe imediat sinteza moleculelor de ADN. Prin urmare, citoplasma celulelor aflate in faza S contine factori care initiaza sinteza de ADN in nucleul celulelor din faza G1. Fuzionarea celulelor aflate in faza G2 cu cele din faza S determina un efect diferit: nucleul G2 este incapabil sa initieze sinteza de ADN, chiar daca este prezenta citoplasma de faza S, care contine factorii respectivi. De aici rezulta ca este prevenita sinteza de ADN in nucleul de faza G2 printr-un mecanism care blocheaza o noua replicare a genomului pana cand se va incheia mitoza.
Mecanisme moleculare de reglare a ciclului celular. Complexe distincte de cicline si kinaze dependente de cicline (Cdk) controleaza trecerea celulelor prin fazele ciclului celular.
Ciclinele sunt subunitati de reglare ale acestor complexe care activeaza proteinkinazele. Au fost identificate pana in prezent cel putin opt cicline, notate de la A la H.
Kinazele dependente de cicline (Cdk) au proprietatea de a fosforila substraturile, activitatea lor fiind reglata prin diferite mecanisme: asocierea cu ciclinele, fosforirilare - defosforilare si interactiunea cu proteine inhibitorii specifice. Din celulele eucariote au fost izolate sapte proteine Cdk.
Spre exemplu:
complexul Cdk4/6 - ciclina D permite celulei sa depaseasca punctul G1;
complexul Cdk 2 - ciclina E intervine in faza tarzie G1si in faza S;
complexul Cdk2 - ciclina A asigura parcurgerea fazei S;
complexul Cdk1 - ciclina A intervine in faza tarzie G2;
complexul Cdk1(Cdc2) - ciclina B asigura tranzitia celulelor eucariote din faza G2 in faza M.
Dupa modul de parcurgere a ciclului celular se cunosc trei categorii de celule:
celule oprite in faza G1, care dupa diferentierea in cursul dezvoltarii embrionare se mentin fara a se mai divide pe toata durata vietii (ex.: miocitele, adipocitele, neuronii, celulele cristalinului);
celule care au o durata de viata scurta si se divid foarte rapid (ex.: enterocitele, celulele stem, celulele liniei seminale);
celule cu capacitate moderata de multiplicare, dar care in anumite conditii se pot divide rapid (ex.: hepatocitele, fibroblastele din piele, celulele endoteliale ale vaselor sangvine, celulele endocrine).
In tesuturile alcatuite din celule cu durata scurta de viata si respectiv multiplicare rapida, exista doua compartimente celulare:
un compartiment neproliferativ;
un compartiment proliferativ care cuprinde celulele stem (celule de rezerva, aflate in faza G0, care, sub influenta factorilor stimulatori ai multiplicarii, trec in compartimentul proliferativ unde se divid rapid, compensand pierderile de celule mature).
Lezarea moleculelor de ADN poate opri ciclul celular si in faza G1. Si in acest caz, stationarea in faza G1 permite repararea leziunilor, inainte ca celulele sa intre in faza S (situatie in care ADN-ul lezat s-ar replica). In celulele de origine animala, oprirea ciclului celular in punctul de restrictie este mediata de actiunea unei proteine p53, care este rapid indusa ca raspuns la lezarea ADN-ului. Interesant de semnalat este faptul ca gena care codifica proteina p53 sufera frecvent mutatii in cancerul uman. In urma aparitiei unor mutatii, pierderea functionalitatii proteinei p53 impiedica oprirea ciclului celular in faza G1 si, astfel, in schimbul repararii, ADN-ul lezat este transmis celulelor fiice. Mostenirea unor molecule de ADN lezat determina o crestere a frecventei mutatiilor si a instabilitatii generale a genomului celular, care contribuie la dezvoltarea proceselor tumorale. Mutatiile din structura genei p53 sunt cele mai frecvente alterari genetice intalnite drept cauza in cancerologia umana, ilustrand importanta reglarii ciclului celular pentru viata organismelor multicelulare.
| Contact |- ia legatura cu noi -| | |
| Adauga document |- pune-ti documente online -| | |
| Termeni & conditii de utilizare |- politica de cookies si de confidentialitate -| | |
| Copyright © |- 2026 - Toate drepturile rezervate -| |
|
|
|||
|
|||
|
|||
Proiecte pe aceeasi tema | |||
|
| |||
|
|||
|
|
|||